



Fattori pre-raccolta per una produzione ottimale di ciliegie dolci
Sviluppo del frutto: acqua, carico produttivo, chioma e nutrizione
La crescita del frutto di ciliegio dolce viene comunemente descritta secondo un andamento doppio sigmoide, organizzato in tre fasi fisiologiche.
La Fase I è dominata dalla divisione cellulare e dalla crescita iniziale del mesocarpo; durante la Fase II, la crescita esterna rallenta mentre proseguono lo sviluppo dell’embrione e l’indurimento dell’endocarpo; nella Fase III, o fase di accrescimento del frutto, riprende un’intensa espansione cellulare, associata all’afflusso di acqua, all’accumulo di zuccheri, allo sviluppo del colore e alla maturazione commerciale.
Questa sequenza spiega perché irrigazione, carico produttivo, ambiente luminoso della chioma e nutrizione non esercitano lo stesso effetto nelle diverse fasi: un intervento utile in una determinata finestra può essere irrilevante o persino controproducente in un’altra (Wani et al., 2014; Vignati et al., 2024).
Durante la fase di accrescimento, la dinamica vascolare del frutto subisce un cambiamento critico.
Gestione dell’acqua
Il flusso idrico xilematico diminuisce progressivamente fino a diventare quasi trascurabile alla maturità, mentre il contributo floematico aumenta e diventa la via dominante per l’ingresso di acqua e assimilati.
Questa transizione riduce la capacità del frutto maturo di compensare brusche variazioni dello stato idrico e contribuisce a spiegare la sua elevata sensibilità alle fluttuazioni idriche durante le fasi finali dello sviluppo (Brüggenwirth et al., 2016).
La gestione dell’acqua durante la maturazione deve essere interpretata a partire da questa fisiologia.
Un grave deficit idrico può limitare la crescita, ridurre la pezzatura dei frutti e compromettere la qualità commerciale; al contrario, un’eccessiva disponibilità d’acqua o l’alternanza brusca tra deficit e reidratazione può creare condizioni predisponenti a microcracking e cracking.
Programmazione dell’irrigazione
Negli agroecosistemi montani, altitudine e programmazione dell’irrigazione condizionano lo stato idrico dell’albero, la fenologia e la pezzatura dei frutti, rafforzando la necessità di adattare la gestione dell’acqua a cultivar, portinnesto, suolo e domanda atmosferica locale (Nieto-Serrano et al., 2026).
Inoltre, il contributo vascolare all’approvvigionamento idrico può partecipare ad alcuni tipi di cracking, soprattutto quando coincidono frutti maturi, elevata suscettibilità varietale ed eventi di pioggia o reidratazione (Measham et al., 2010; Knoche et al., 2025).
La programmazione dell’irrigazione dovrebbe quindi essere collegata allo stadio fenologico, alla riserva idrica del suolo, alla domanda evaporativa, allo stato idrico dell’albero e alle previsioni di pioggia.
Nella pratica, il rischio maggiore spesso non è rappresentato da un singolo livello di disponibilità idrica, ma da una sequenza di stress e rapida reidratazione durante il periodo di accrescimento finale e maturazione.
Carico produttivo
Il monitoraggio dovrebbe mirare a ridurre le brusche oscillazioni fisiologiche, mantenendo al tempo stesso un apporto idrico sufficiente per la crescita dei frutti e la funzionalità della chioma.
Carico produttivo e architettura della chioma condizionano le relazioni sorgente-pozzo e il microclima dei frutti.
Un carico produttivo elevato aumenta la competizione tra frutti, può ridurre la pezzatura e può modificare l’accumulo di solidi solubili; un carico produttivo eccessivamente basso, tuttavia, può favorire vigore vegetativo, ombreggiamento e squilibri nella distribuzione delle risorse.
Potatura, forma di allevamento e regolazione del carico produttivo devono essere calibrate in base a cultivar, portinnesto, vigore, disponibilità idrica e obiettivo commerciale, senza estendere automaticamente a tutti i frutteti risposte ottenute in un solo sistema di allevamento, a una sola altitudine o sotto una specifica infrastruttura di protezione.
Equilibrio sorgente-pozzo
L’obiettivo rilevante non è la riduzione del carico in sé, ma un equilibrio funzionale tra numero di frutti, area fogliare, intercettazione luminosa e disponibilità di assimilati (Whiting & Ophardt, 2005; Wani et al., 2014; Blanco et al., 2019; Penzel et al., 2020; Nieto-Serrano et al., 2026).
Questa interpretazione sorgente-pozzo ha conseguenze dirette sulla gestione.
La domanda giornaliera stimata di area fogliare per frutto aumenta durante lo sviluppo e può raggiungere valori elevati durante la Fase III, con stime specifiche per cultivar riportate per 'Bellise' e 'Regina'.
Regolazione del carico produttivo, diradamento dei fiori e adeguamento del carico di frutti possono migliorare la pezzatura e alcuni attributi qualitativi in determinati sistemi, ma le risposte non sono universali e possono variare in funzione di cultivar, portinnesto, densità di fruttificazione, capacità della chioma, sistema di copertura e struttura dei prezzi di mercato.
Nutrizione minerale
Le decisioni sul carico produttivo dovrebbero quindi essere prese in funzione di un obiettivo commerciale definito, e non come prescrizione generica di diradamento (Whiting & Ophardt, 2005; Einhorn et al., 2011; Penzel et al., 2020; González-Villagra et al., 2024; Parveze et al., 2024).
La nutrizione minerale dovrebbe essere interpretata in modo funzionale in base allo stadio fenologico.
Il potassio partecipa al trasporto degli zuccheri, alla regolazione osmotica e alla qualità durante l’accrescimento del frutto, rendendo la sua disponibilità particolarmente rilevante durante la Fase III.
Il calcio contribuisce all’integrità della parete cellulare e alla riduzione del rigonfiamento della parete cellulare epidermica, sebbene la sua bassa mobilità verso il frutto e il dilavamento causato dalla pioggia limitino l’efficacia delle applicazioni tardive.
Risposta ai nutrienti
Le risposte ai trattamenti fogliari devono quindi essere interpretate in base a formulazione, tempistica, copertura, cultivar, cuticola, condizioni ambientali e reale rischio di bagnatura (Wani et al., 2014; Schumann et al., 2022; Kafle et al., 2016; Varaldo & Giacalone, 2025a).
La Tabella 5 riassume i nutrienti più rilevanti per lo sviluppo del frutto e la qualità.
La relazione tra nutrizione, consistenza e cracking deve essere formulata con cautela.
I trattamenti fogliari a base di calcio e potassio possono migliorare le proprietà meccaniche e alcuni attributi qualitativi in specifiche condizioni sperimentali, ma non dovrebbero essere presentati come una soluzione universale al cracking.
Nutrizione e cracking
Il loro effetto dipende da cultivar, stadio di maturazione, suscettibilità cuticolare, microcracking pregresso, gestione dell’acqua, formulazione, dilavamento da pioggia e reale pressione di bagnatura.
In questa sezione, la nutrizione viene quindi affrontata come supporto funzionale alla crescita e alla qualità; la fisiologia specifica del cracking indotto dalla pioggia viene sviluppata nella Sezione 8 (Schumann et al., 2022; Kafle et al., 2016; Knoche et al., 2025; Varaldo & Giacalone, 2025a).
Tabella 5. Nutrienti chiave per stadio fenologico e funzione produttiva
| Nutriente | Finestra critica | Funzione principale | Rischio gestionale | Fonti principali |
|---|---|---|---|---|
| Azoto (N) | Post-raccolta, germogliamento e crescita iniziale. | Area fogliare, riserve e capacità fotosintetica. | Eccesso: vigore, ombreggiamento, frutti più molli e minore differenziazione fiorale. | Michailidis et al. (2018); Wani et al. (2014) |
| Boro (B) | Pre-fioritura, fioritura e allegagione. | Polline, tubetto pollinico e parete cellulare. | Margine ristretto tra carenza e tossicità; è necessaria la diagnosi. | Hedhly et al. (2004); Xu et al. (2023) |
| Zinco (Zn) | Ingrossamento delle gemme e germogliamento. | Crescita iniziale, attività enzimatica e fertilità delle gemme. | Le applicazioni richiedono validazione locale. | Michailidis et al. (2018); Wani et al. (2014) |
| Potassio (K) | Accrescimento del frutto e maturazione. | Trasporto degli zuccheri, regolazione osmotica e qualità. | Squilibrio con Ca/Mg o aumento del turgore in condizioni di cattiva gestione idrica. | Wani et al. (2014); Varaldo & Giacalone (2025a) |
| Calcio (Ca) | Allegagione, divisione cellulare e maturazione. | Parete cellulare, consistenza e minore rigonfiamento epidermico. | Bassa mobilità, dilavamento da pioggia e risposta fogliare variabile; non è una soluzione autonoma. | Schumann et al. (2022); Kafle et al. (2016); Varaldo & Giacalone (2025a) |
Nota: La gestione dei nutrienti dovrebbe essere supportata da analisi fogliari e del suolo, diagnosi locale, cultivar, portinnesto e normative vigenti.
Maturazione, piogge, cracking e sistemi di produzione protetta
La maturazione concentra valore commerciale e vulnerabilità.
In questa fase finale, il frutto esprime colore, pezzatura, consistenza, solidi solubili, acidità e sapore, ma diventa anche più sensibile a pioggia, grandine, bagnatura superficiale prolungata, cracking, microcracking e marciumi associati.
L’incidenza del cracking e la sua tipologia variano in funzione di genotipo, stagione, stadio di maturazione e condizioni ambientali durante l’evento piovoso, spiegando perché la stessa precipitazione può generare risposte molto diverse tra cultivar, appezzamenti o annate (Measham et al., 2009; Blanco et al., 2019; Rojas et al., 2021).
Il cracking indotto dalla pioggia non dovrebbe essere interpretato come la semplice conseguenza di un massiccio assorbimento d’acqua e di aumenti uniformi del turgore del frutto.
Bagnatura superficiale
Le evidenze sperimentali disponibili indicano che la bagnatura superficiale localizzata è un fattore scatenante critico: l’acqua trattenuta sulla buccia penetra preferenzialmente attraverso microcracking e discontinuità cuticolari, favorendo danni localizzati nei tessuti epidermici e ipodermici che possono propagarsi in spaccature visibili (Winkler et al., 2020; Knoche et al., 2022).
Il contributo vascolare può essere rilevante per specifici tipi di spacco, in particolare il cracking laterale, ma dovrebbe essere formulato come via complementare e dipendente dal contesto, non come unica spiegazione del fenomeno (Measham et al., 2010).
La suscettibilità al cracking integra proprietà genetiche, stadio di maturazione, pezzatura e consistenza del frutto, caratteristiche della cuticola, microcracking pregresso, calcio nei tessuti e durata della bagnatura superficiale.
Questa integrazione collega la variabilità osservata tra genotipi e stagioni con i meccanismi locali di perdita dell’integrità della buccia.
Modello della cerniera lampo
In famiglie di pieni fratelli contrastanti per suscettibilità al cracking, la tolleranza è stata associata a caratteri cuticolari, microcracking e calcio, rafforzando la necessità di interpretare il cracking come un problema di struttura della buccia, più che come una semplice risposta generale del frutto all’acqua (Measham et al., 2009; Knoche et al., 2025).
Il modello della “cerniera lampo” fornisce un quadro meccanicistico per spiegare la propagazione del danno.
Secondo questo modello, la bagnatura superficiale e il microcracking facilitano l’ingresso localizzato di acqua, la lisi delle cellule epidermiche o subepidermiche e il rilascio di acidi organici, soprattutto malato, nell’apoplasto.
Questo processo può estrarre calcio dalle pareti cellulari vicine, aumentare il rigonfiamento della parete, ridurre l’adesione cellulare e favorire la progressiva scucitura della buccia.
Sequenza meccanicistica
Il modello non dovrebbe essere trattato come una spiegazione chiusa per tutte le cultivar e condizioni, ma come un’ipotesi integrativa che collega cuticola, acqua superficiale, calcio, parete cellulare e propagazione dello spacco (Knoche & Winkler, 2019; Schumann et al., 2022; Knoche et al., 2022).
La Figura 4 sintetizza questa sequenza meccanicistica e la sua interpretazione applicata.
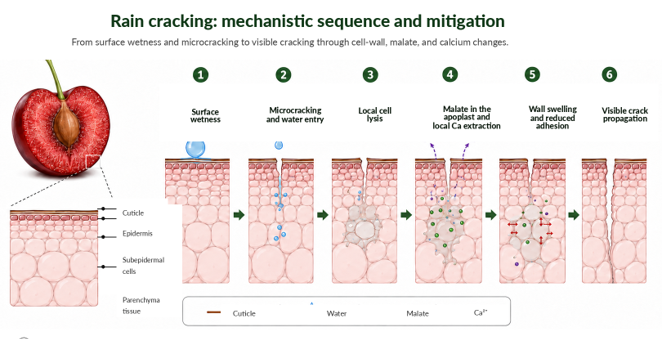
Figura 4. Sequenza meccanicistica proposta per il cracking indotto dalla pioggia nel ciliegio dolce.
Lo schema sintetizza la transizione dalla bagnatura superficiale e dal microcracking cuticolare all’ingresso localizzato di acqua, alla lisi cellulare, al rilascio di malato nell’apoplasto, all’estrazione locale di calcio, al rigonfiamento della parete, alla perdita di adesione cellulare e alla propagazione visibile dello spacco.
Strategie di mitigazione
Le strategie di mitigazione devono agire su più punti del sistema: ridurre la durata della bagnatura superficiale del frutto, evitare brusche oscillazioni dello stato idrico, mantenere una chioma ventilata, diminuire l’esposizione diretta a pioggia o grandine e applicare programmi nutrizionali o trattamenti pre-raccolta solo quando diagnosi ed evidenze locali li supportano.
Le applicazioni fogliari di calcio o potassio possono migliorare le proprietà meccaniche e ridurre la suscettibilità al cracking in prove specifiche, ma la loro efficacia dipende da formulazione, dose, tempistica, copertura, dilavamento da pioggia, cultivar e stadio di maturazione (Kafle et al., 2016; Schumann et al., 2022; Varaldo & Giacalone, 2025a).
Analogamente, i trattamenti con acido abscissico, metil jasmonato o melatonina hanno mostrato effetti favorevoli in contesti specifici, anche se dovrebbero essere considerati strategie condizionate da cultivar, stadio fenologico, dose, stato normativo e reale pressione pluviometrica (Balbontín et al., 2018; Ruiz-Aracil et al., 2024).
Poiché la bagnatura superficiale è centrale nel rischio di cracking, le tecnologie di monitoraggio possono aggiungere valore quando stimano la bagnatura del frutto, e non solo la pioggia.
Sistemi protetti
Le immagini termiche-RGB combinate con sensori meteorologici in campo hanno prodotto modelli di previsione della bagnatura specifici per cultivar, mostrando come temperatura superficiale del frutto e microclima locale possano essere integrati nel supporto decisionale per asciugatura, trattamenti o strategie di protezione dalla pioggia (Ranjan et al., 2022).
Coperture plastiche, tunnel alti e altri sistemi protetti riducono il contatto diretto tra pioggia e frutto e possono diminuire le perdite da cracking o grandine.
Tuttavia, non sono strutture neutre: modificano radiazione, temperatura, umidità relativa, ventilazione, vigore, maturazione, qualità e gestione delle malattie.
I tunnel alti hanno dimostrato di proteggere dalla pioggia e di modificare variabili fisiologiche e produttive, mentre i sistemi di forzatura sotto copertura possono alterare radiazione, temperatura, consistenza, solidi solubili e composti bioattivi.
Sistema produttivo
Studi recenti in condizioni mediterranee hanno inoltre mostrato che i sistemi con copertura a tetto e tunnel alto possono ridurre la radiazione incidente, modificare il microclima a livello della chioma e lo stato idrico dell’albero, anticipare la raccolta e influire su colore, consistenza, acidità, fenoli e capacità antiossidante.
La protezione fisica dovrebbe quindi essere valutata come una modifica del sistema produttivo e come investimento di gestione del rischio, non semplicemente come una barriera contro un evento meteorologico isolato (Schmitz-Eiberger & Blanke, 2012; Blanco et al., 2019; Rojas et al., 2021; Palma et al., 2023).
Cambiamento climatico e adattamento del sistema produttivo
Il cambiamento climatico modifica il quadro agroclimatico su cui si fonda la produzione di ciliegio dolce, perché influisce contemporaneamente sull’accumulo di freddo invernale, sul calore primaverile, sulla sincronia fenologica e sulla frequenza degli eventi estremi durante fioritura, allegagione e maturazione.
Nel ciliegio dolce, il riscaldamento non implica necessariamente un semplice anticipo fenologico.
Rischio di dormienza
Quando il fabbisogno di freddo non viene soddisfatto, il rilascio dell’endodormienza può essere incompleto, con germogliamento irregolare, fioritura disomogenea e minore stabilità produttiva.
Al contrario, quando il fabbisogno di freddo è soddisfatto, una primavera più calda può accelerare l’accumulo di calore e anticipare la fioritura, aumentando l’esposizione degli organi riproduttivi alle gelate tardive.
L’adattamento non può quindi basarsi su un singolo indicatore termico; deve considerare l’interazione tra freddo, forcing, cultivar, data di fioritura e rischio fenologico locale (Fadón et al., 2021; Fadón et al., 2023).
La vulnerabilità climatica è fortemente specifica per cultivar.
Idoneità delle cultivar
Gli studi sui requisiti agroclimatici nelle cultivar spagnole e commerciali mostrano che i genotipi differiscono per fabbisogno di freddo e calore e per capacità di mantenere una fioritura regolare in scenari di riscaldamento.
Le proiezioni in condizioni mediterraneo-continentali nella Valle dell’Ebro indicano che, entro il 2050, la maggior parte delle cultivar valutate potrebbe ancora mantenere una ragionevole idoneità climatica, sebbene le cultivar con fabbisogni di freddo più elevati mostrino rischi più precoci di disadattamento.
Verso la fine del secolo, soprattutto in scenari ad alte emissioni, anche cultivar a basso fabbisogno di freddo potrebbero rischiare di non soddisfare le proprie esigenze di freddo in alcuni modelli climatici.
Questi risultati non dovrebbero essere estrapolati senza una calibrazione regionale, ma sottolineano che la selezione varietale e la zonazione agroclimatica diventeranno decisioni sempre più critiche (Fadón et al., 2023; Santolaria et al., 2026).
Adattamento strutturale
L’adattamento strutturale inizia prima dell’impianto.
Comprende sito, altitudine, esposizione, cultivar, portinnesto, disponibilità idrica, forma di allevamento e infrastrutture di protezione.
In agroecosistemi montani come la Valle del Jerte, le misurazioni fisiologiche e meteorologiche mostrano che una maggiore altitudine può ridurre la domanda evaporativa, migliorare lo stato idrico dell’albero, ritardare la fenologia ed estendere il periodo di sviluppo del frutto, favorendo pezzature commerciali maggiori rispetto alle aree più basse esposte a una combinazione più intensa di calore e stress idrico.
Questa evidenza supporta il valore dell’altitudine come fattore locale di adattamento, anche se il suo impiego deve essere interpretato all’interno di ciascun sistema produttivo e dei relativi limiti in termini di suolo, pendenza, acqua, accessibilità e rischio di gelo (Nieto-Serrano et al., 2026).
Infrastrutture di protezione
Le infrastrutture di protezione fanno parte dell’adattamento strutturale, ma non dovrebbero essere considerate neutre.
I tunnel alti e le coperture plastiche possono ridurre le perdite da pioggia o grandine e modificare la data di maturazione, ma alterano anche radiazione, temperatura, umidità relativa, ventilazione, vigore, qualità e gestione delle malattie.
La loro utilità dipende da rischio climatico, cultivar, sistema di allevamento, costi di installazione, logistica gestionale e obiettivo di mercato.
Le decisioni di investimento in coperture o altre tecnologie di protezione dovrebbero quindi essere valutate come problemi di gestione del rischio, non semplicemente come risposte episodiche a un evento meteorologico (Blanco et al., 2019; Rojas et al., 2021).
Misure operative
Accanto alle misure strutturali, l’adattamento richiede misure operative adeguate a ogni stagione.
Tra queste rientrano il monitoraggio orario del freddo e del calore, il tracciamento fenologico, la gestione dell’irrigazione, l’adeguamento del carico produttivo, la potatura per modulare il microclima della chioma e la preparazione a gelate, pioggia o grandine.
Irrigazione, carico produttivo e gestione della chioma possono contribuire a modulare lo stato idrico dell’albero, il microclima dei frutti e l’esposizione ai rischi della maturazione finale, mentre irrigazione in deficit controllato e gestione dell’acqua devono sempre essere formulate in base a cultivar, portinnesto, suolo, fase fenologica e disponibilità idrica.
L’adattamento operativo non è quindi l’applicazione di ricette fisse, ma la riduzione delle oscillazioni fisiologiche che aumentano la vulnerabilità del frutto e dell’albero (Blanco et al., 2019; Nieto-Serrano et al., 2026).
Supporto decisionale
In questo quadro, il monitoraggio strumentale e i sistemi di supporto decisionale possono aiutare a tradurre la comprensione fisiologica in decisioni operative più precise.
L’integrazione di dati orari su freddo, calore, pioggia, umidità, temperatura di bulbo umido, bagnatura superficiale del frutto, monitoraggio fenologico, indicatori dello stato idrico dell’albero e valutazione probabilistica del rischio può guidare irrigazione, protezione dal gelo, previsione della bagnatura superficiale associata al cracking e investimento in coperture o reti in condizioni locali.
Questi strumenti non sostituiscono il giudizio agronomico né eliminano l’incertezza, ma migliorano la tracciabilità delle decisioni quando vengono interpretati insieme a cultivar, portinnesto, microclima, maturità del frutto, infrastrutture disponibili e obiettivo commerciale (Fadón et al., 2021; Fadón et al., 2023; Rojas et al., 2021; Ranjan et al., 2022; Nieto-Serrano et al., 2026).
Nel complesso, l’adattamento del ciliegio dolce al cambiamento climatico richiede l’integrazione di modelli termici, fisiologia dell’albero, rischio economico e gestione localizzata.
Strategia di adattamento
Le strategie più solide combineranno zonazione agroclimatica delle cultivar, selezione di cultivar e portinnesti adattati, protezione fisica quando giustificata, gestione efficiente dell’acqua, monitoraggio fenologico e analisi economica delle decisioni.
Questa interpretazione chiude il capitolo con un’idea centrale: la produzione ottimale di ciliegie dolci dipenderà non solo dall’aumento della resa o della qualità in condizioni medie, ma dalla progettazione di sistemi capaci di mantenere stabilità produttiva in presenza di inverni più irregolari, primavere più incerte ed eventi estremi più frequenti (Rojas et al., 2021; Fadón et al., 2023; Santolaria et al., 2026).
La Tabella 6 riassume i principali fattori pre-raccolta, la loro tempistica critica, l’effetto produttivo atteso e il relativo orientamento tecnico.
La sua interpretazione deve rimanere contestuale e tracciabile rispetto alle evidenze disponibili.
Matrice pre-raccolta
Tabella 6. Matrice sintetica dei fattori pre-raccolta e orientamento gestionale contestualizzato
| Tempistica | Fattore critico | Effetto produttivo | Orientamento tecnico |
|---|---|---|---|
| Impianto | Sito, altitudine, esposizione, drenaggio, cultivar, portinnesto e infrastruttura. | Definisce freddo, gelo, vigore, acqua, salute radicale ed esposizione agli eventi estremi. | Dare priorità all’adattamento strutturale locale e non separare la cultivar da portinnesto, microclima e disponibilità idrica. |
| Ciclo precedente | Calore estivo, riserve, vigore e nutrizione. | Condiziona induzione fiorale e qualità delle gemme. | Evitare stress severi e vigore eccessivo; mantenere una chioma illuminata. |
| Dormienza-germogliamento | Freddo CP/CH/CU e forcing GDH. | Regola uniformità e data di fioritura. | Usare dati orari e modelli espliciti; evitare soglie universali. |
| Fioritura-allegagione | Gelo, compatibilità, EPP e impollinatori. | Determina fecondazione, allegagione e carico produttivo iniziale. | Progettare impollinatori compatibili e attivare la mitigazione in base alla temperatura di bulbo umido e alla fenofase. |
| Accrescimento-maturazione | Acqua, domanda evaporativa, carico produttivo, equilibrio K-Ca, pioggia, copertura e microclima. | Definisce pezzatura, qualità, consistenza, cracking e perdite finali. | Adeguare irrigazione, carico produttivo, chioma e nutrizione; valutare la protezione fisica in base a rischio e costo. |
| Adattamento climatico | Freddo, forcing, altitudine, acqua, gelo, pioggia e grandine. | Condiziona idoneità varietale, stabilità fenologica e rischio economico. | Combinare zonazione, monitoraggio orario, gestione operativa e analisi decisionale. |
Nota: Questa matrice è concepita come quadro di decisione contestuale e non dovrebbe essere usata come prescrizione fissa.
Dal punto di vista operativo, la matrice è utilizzata al meglio come ciclo di monitoraggio stagionale: prima si definisce il rischio strutturale del sito e della cultivar, poi si aggiorna l’accumulo di freddo e forcing durante la dormienza, si intensificano le decisioni su gelo e impollinazione durante la fioritura, si adeguano acqua e carico produttivo durante lo sviluppo del frutto e si rivalutano pioggia, bagnatura, cracking e rischio di raccolta durante la maturazione.
Questa sequenza trasforma la catena fisiologica descritta nel capitolo in un processo di gestione tracciabile.
Conclusioni
La produzione ottimale di ciliegie dolci dovrebbe essere compresa come una catena fisiologica e agronomica continua, in cui la resa commerciale di una stagione dipende da processi avviati durante il ciclo precedente.
Induzione fiorale, qualità delle gemme, accumulo di freddo invernale, calore primaverile, sensibilità al gelo, fioritura, impollinazione e allegagione operano come stadi interdipendenti.
Resa commerciale
Il successo produttivo non può quindi essere attribuito a una singola pratica gestionale o a un singolo indicatore termico, ma alla coerenza tra cultivar, portinnesto, sito, fenologia, microclima e decisioni agronomiche adattate a ciascun sistema.
Durante lo sviluppo del frutto, la gestione di acqua, carico produttivo, chioma e nutrizione minerale condiziona l’espressione finale di pezzatura, consistenza, colore, solidi solubili e qualità commerciale.
La maturazione, tuttavia, concentra sia il valore più elevato sia la maggiore vulnerabilità della coltura.
In questa fase, la riduzione del contributo xilematico, l’afflusso dominato dal floema, l’accumulo di soluti, la tensione cuticolare e il microcracking aumentano la sensibilità del frutto a pioggia, bagnatura superficiale, cracking, grandine e marciumi.
Rischi finali
La qualità costruita nel corso di mesi può essere persa rapidamente se i rischi della maturazione finale non vengono anticipati attraverso gestione dell’acqua, protezione fisica, ventilazione della chioma, nutrizione contestualizzata e decisioni di raccolta adeguate.
Nel contesto del cambiamento climatico, la stabilità produttiva del ciliegio dolce dipenderà sempre più dalla capacità di adattare i sistemi frutticoli a inverni più irregolari, primavere più incerte, maggiore combinazione di calore e stress idrico ed eventi estremi più frequenti.
Questo adattamento dovrebbe essere organizzato su due livelli complementari: misure strutturali, come scelta della regione, altitudine, cultivar, portinnesto, disegno d’impianto e infrastrutture di protezione; e misure operative, come monitoraggio fenologico, adeguamento dell’irrigazione, regolazione del carico produttivo, gestione della chioma e risposta a gelo, pioggia o grandine.
Nessuna di queste misure dovrebbe essere interpretata come una ricetta universale, ma come parte di una strategia locale basata su diagnosi, evidenze e analisi del rischio.
Approccio di sistema
Nel complesso, la gestione pre-raccolta del ciliegio dolce dovrebbe passare da interventi isolati a un approccio di sistema.
Una produzione commerciale stabile si ottiene non solo massimizzando resa o qualità in condizioni medie, ma riducendo la vulnerabilità dell’albero e del frutto durante le finestre fenologiche più sensibili.
L’integrazione di fisiologia, dati agroclimatici, soglie specifiche per cultivar, monitoraggio locale e analisi decisionale sarà essenziale per convertire il potenziale biologico del ciliegio dolce in frutti di alta qualità, produzione resiliente e resa commerciale sostenibile.
Jesús Alonso¹,²
¹ Institute of Food Science, Technology and Nutrition (ICTAN-CSIC), Spanish National Research Council (CSIC), c/ José Antonio Novais, 6, 28040 Madrid, Spain.
² University Research Institute for Agricultural Resources (INURA), Universidad de Extremadura, Edificio de los Institutos Universitarios de Investigación, Avenida de la Investigación s/n, 06006 Badajoz, Spain.
Riferimenti bibliografici
- Alburquerque, N., García-Montiel, F., Carrillo, A., Burgos, L., 2008. Chilling and heat requirements of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements. Environmental and Experimental Botany, 64(2), 162-170. https://doi.org/10.1016/j.envexpbot.2008.01.003
- Ayala, M., Lang, G.A., 2015. 13C-photoassimilate partitioning in sweet cherry (Prunus avium) during early spring. Ciencia e Investigación Agraria, 42(2), 191-203. https://doi.org/10.4067/S0718-16202015000200006
- Azizi Gannouni, T., Campoy, J.A., Quero-García, J., Barreneche, T., Arif, A., Albouchi, A., Ammari, Y., 2017. Dormancy related traits and adaptation of sweet cherry in Northern Africa: A case of study in two Tunisian areas. Scientia Horticulturae, 219, 272-279. https://doi.org/10.1016/j.scienta.2017.03.013
- Balbontín, C., Gutiérrez, C., Wolff, M., Figueroa, C.R., 2018. Effect of abscisic acid and methyl jasmonate preharvest applications on fruit quality and cracking tolerance of sweet cherry. Chilean Journal of Agricultural Research, 78(3), 438-446. https://doi.org/10.4067/S0718-58392018000300438
- Blanco, V., Zoffoli, J.P., Ayala, M., 2019. High tunnel cultivation of sweet cherry (Prunus avium L.): Physiological and production variables. Scientia Horticulturae, 251, 108-117. https://doi.org/10.1016/j.scienta.2019.02.023
- Bonomelli, C., Bonilla, C., Acuña, E., Artacho, P., 2012. Seasonal pattern of root growth in relation to shoot phenology and soil temperature in sweet cherry trees (Prunus avium): A preliminary study in central Chile. Ciencia e Investigación Agraria, 39(1), 127-136.
- Brüggenwirth, M., Winkler, A., Knoche, M., 2016. Xylem, phloem, and transpiration flows in developing sweet cherry fruit. Trees, 30, 1821-1830. https://doi.org/10.1007/s00468-016-1415-4
- Choi, C., Tao, R., Andersen, R.L., 2002. Identification of self-incompatibility alleles and pollen incompatibility groups in sweet cherry by PCR-based S-allele typing and controlled pollination. Euphytica, 123, 9-20. https://doi.org/10.1023/A:1014403802677
- Djordjevic, B.S., Djurovic, D.B., Zec, G.N., Boskov, D.D., 2021. The influence of rootstocks on the sensitivity of flower buds to frost and the main properties of the 'Carmen' sweet cherry cultivar. Journal of Agricultural Sciences (Belgrade), 66(4), 351-358. https://doi.org/10.2298/JAS2104351D
- Dziedzic, E., Bieniasz, M., Kowalczyk, B., 2019. Morphological and physiological features of sweet cherry floral organ affecting the potential fruit crop in relation to the rootstock. Scientia Horticulturae, 251, 127-135. https://doi.org/10.1016/j.scienta.2019.03.013
- Eeraerts, M., Meeus, I., Van Den Berge, S., Smagghe, G., 2017. Landscapes with high intensive fruit cultivation reduce wild pollinator services to sweet cherry. Agriculture, Ecosystems & Environment, 239, 342-348. https://doi.org/10.1016/j.agee.2017.01.031
- Eeraerts, M., Smagghe, G., Meeus, I., 2020a. Bumble bee abundance and richness improves honey bee pollination behaviour in sweet cherry. Basic and Applied Ecology, 43, 27-33. https://doi.org/10.1016/j.baae.2019.11.004
- Eeraerts, M., Vanderhaegen, R., Smagghe, G., Meeus, I., 2020b. Pollination efficiency and foraging behaviour of honey bees and non-Apis bees to sweet cherry. Agricultural and Forest Entomology, 22, 75-82. https://doi.org/10.1111/afe.12363
- Einhorn, T.C., Laraway, D., Turner, J., 2011. Crop load management does not consistently improve crop value of 'Sweetheart'/'Mazzard' sweet cherry trees. HortTechnology, 21(5), 546-553.
- Fadón, E., Fernández, E., Luedeling, E., Rodrigo, J., 2023. Agroclimatic requirements and adaptation potential to global warming of Spanish cultivars of sweet cherry (Prunus avium L.). European Journal of Agronomy, 145, 126774. https://doi.org/10.1016/j.eja.2023.126774
- Fadón, E., Herrero, M., Rodrigo, J., 2015. Flower development in sweet cherry framed in the BBCH scale. Scientia Horticulturae, 192, 141-147. https://doi.org/10.1016/j.scienta.2015.05.027
- Fadón, E., Herrero, M., Rodrigo, J., 2018a. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science, 9, 171. https://doi.org/10.3389/fpls.2018.00171
- Fadón, E., Rodrigo, J., Herrero, M., 2018b. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees, 32(6), 1583-1594. https://doi.org/10.1007/s00468-018-1735-7
- Fadón, E., Rodrigo, J., Luedeling, E., 2021. Cultivar-specific responses of sweet cherry flowering to rising temperatures during dormancy. Agricultural and Forest Meteorology, 307, 108486. https://doi.org/10.1016/j.agrformet.2021.108486
- Feldmane, D., Sebre, G., Rubauskis, E., 2025. Cultivar and rootstock effect on sweet cherry fruit density and fruit drop. Rural Sustainability Research, 54(349), 79-86. https://doi.org/10.2478/plua-2025-0019
- García, C.B., Díaz-Siefer, P., Smith-Ramírez, C., Montero-Silva, F., Martínez-Harms, J., Murúa, M., Celis-Diez, J.L., 2025. Synergistic effect of honeybees and wild floral visitors in promoting sweet cherry fruit set in central Chile. Biological Research, 58(1), 39. https://doi.org/10.1186/s40659-025-00617-2
- González-Villagra, J., Palacios-Peralta, C., Muñoz-Alarcón, A., Reyes-Díaz, M., Osorio, P., Ribera-Fonseca, A., 2024. Influence of fruit load regulation on harvest and postharvest fruit quality and antioxidant-related parameters in sweet cherry (Prunus avium L.) cv. Regina cultivated under plastic covers in Southern Chile. Plants, 13, 2257. https://doi.org/10.3390/plants13162257
- Götz, K.-P., Naher, J., Fettke, J., Chmielewski, F.-M., 2018. Changes of proteins during dormancy and bud development of sweet cherry (Prunus avium L.). Scientia Horticulturae, 239, 41-49. https://doi.org/10.1016/j.scienta.2018.05.016
- Hedhly, A., Hormaza, J.I., Herrero, M., 2004. Effect of temperature on pollen tube kinetics and dynamics in sweet cherry, Prunus avium (Rosaceae). American Journal of Botany, 91(4), 558-564. https://doi.org/10.3732/ajb.91.4.558
- Hedhly, A., Hormaza, J.I., Herrero, M., 2007. Warm temperatures at bloom reduce fruit set in sweet cherry. Journal of Applied Botany and Food Quality, 81, 158-164. https://doi.org/10.5073/JABFQ.2007.081.025
- Hedhly, A., Hormaza, J.I., Herrero, M., 2009. Flower emasculation accelerates ovule degeneration and reduces fruit set in sweet cherry. Scientia Horticulturae, 119(4), 455-457. https://doi.org/10.1016/j.scienta.2008.08.020
- Holzschuh, A., Dudenhöffer, J.-H., Tscharntke, T., 2012. Landscapes with wild bee habitats enhance pollination, fruit set and yield of sweet cherry. Biological Conservation, 153, 101-107. https://doi.org/10.1016/j.biocon.2012.04.032
- Houghton, E., Watanabe, Y., Neilsen, D., Nelson, L.M., Hannam, K., 2024. Investigating properties of sweet cherry (Prunus avium) flower buds that help promote freezing avoidance by supercooling. Plant Biology, 26(6), 1067-1078. https://doi.org/10.1111/plb.13697
- Kafle, G.K., Khot, L.R., Zhou, J., Bahlol, H.Y., Si, Y., 2016. Towards precision spray applications to prevent rain-induced sweet cherry cracking: Understanding calcium washout due to rain and fruit cracking susceptibility. Scientia Horticulturae, 203, 152-157. https://doi.org/10.1016/j.scienta.2016.03.027
- Karbassioon, A., Yearsley, J., Dirilgen, T., Hodge, S., Stout, J.C., Stanley, D.A., 2023. Responses in honeybee and bumblebee activity to changes in weather conditions. Oecologia, 201, 689-701. https://doi.org/10.1007/s00442-023-05332-x
- Kaufmann, H., Blanke, M.M., 2017. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology, 218, 1-5. https://doi.org/10.1016/j.jplph.2017.07.004
- Kaufmann, H., Blanke, M.M., 2019. Substitution of winter chilling by spring forcing for flowering using sweet cherry as model crop. Scientia Horticulturae, 244, 75-81. https://doi.org/10.1016/j.scienta.2018.09.021
- Kaya, O., Köse, C., 2022. How sensitive are the flower parts of the sweet cherry in sub-zero temperatures? Use of differential thermal analysis and critical temperatures assessment. New Zealand Journal of Crop and Horticultural Science, 50(1), 17-31. https://doi.org/10.1080/01140671.2021.1890156
- Kaya, O., Köse, C., Şahin, M., 2021. The use of differential thermal analysis in determining the critical temperatures of sweet cherry (Prunus avium L.) flower buds at different stages of bud burst. International Journal of Biometeorology, 65(7), 1125-1135. https://doi.org/10.1007/s00484-021-02093-1
- Knoche, M., Grosset-Grange, L., Quero-García, J., Alletru, D., Boutaleb, L., 2025. Cracking susceptibility of full-sibs of a cross of a cracking tolerant and cracking susceptible sweet cherry: Relation to cuticle characteristics, microcracking and calcium. PLOS ONE, 20(1), e0316637. https://doi.org/10.1371/journal.pone.0316637
- Knoche, M., Winkler, A., 2019. The mechanism of rain cracking of sweet cherry fruit. Italus Hortus, 26, 59-65. https://doi.org/10.26353/j.itahort/2019.1.5965
- Knoche, M., Winkler, A., Lang, A., 2022. The unzipping of sweet cherry fruit skin and strategies to prevent it. Italus Hortus, 29(1), 1-13. https://doi.org/10.26353/j.itahort/2022.1.C1
- Laterza, I., Ragone, G., Grossi, G., Cappellari, A., Agostinacchio, M.F., Seclì, G.A., Bari, G., Porro, R., Addante, R., Cornara, D., De Lillo, E., Tamburini, G., 2025. Agricultural intensification at local and landscape scales impacts sweet cherry production through altered pollination services. Agriculture, Ecosystems & Environment, 386, 109586. https://doi.org/10.1016/j.agee.2025.109586
- Mateos-Fierro, Z., Garratt, M.P.D., Fountain, M.T., Ashbrook, K., Westbury, D.B., 2025. Contribution of pollinators to delivering fruit quality in commercial sweet cherry orchards. JSFA Reports. https://doi.org/10.1002/jsf2.70012
- Matzneller, P., Götz, K.-P., Chmielewski, F.-M., 2016. Spring frost vulnerability of sweet cherries under controlled conditions. International Journal of Biometeorology, 60(1), 123-130. https://doi.org/10.1007/s00484-015-1010-1
- McCabe, L.M., Boyle, N.K., Pitts-Singer, T.L., 2024. Osmia lignaria (Hymenoptera: Megachilidae) increase pollination of Washington sweet cherry and pear orchards. Environmental Entomology, 53(4), 698-705. https://doi.org/10.1093/ee/nvae043
- Measham, P.F., Bound, S.A., Gracie, A.J., Wilson, S.J., 2009. Incidence and type of cracking in sweet cherry (Prunus avium L.) are affected by genotype and season. Crop & Pasture Science, 60(10), 1002-1008. https://doi.org/10.1071/CP08410
- Measham, P.F., Gracie, A.J., Wilson, S.J., Bound, S.A., 2010. Vascular flow of water induces side cracking in sweet cherry (Prunus avium L.). Advances in Horticultural Science, 24(4), 243-248. https://www.jstor.org/stable/42883522
- Michailidis, M., Karagiannis, E., Tanou, G., Sarrou, E., Adamakis, I.-D., Karamanoli, K., Martens, S., Molassiotis, A., 2018. Metabolic mechanisms underpinning vegetative bud dormancy release and shoot development in sweet cherry. Environmental and Experimental Botany, 155, 1-11. https://doi.org/10.1016/j.envexpbot.2018.06.024
- Naor, A., Stern, R., Peres, M., Greenblat, Y., Gal, Y., 2005. Timing and severity of postharvest water stress affect following-year productivity and fruit quality of field-grown 'Snow Queen' nectarine. Journal of the American Society for Horticultural Science, 130(6), 806-812. https://doi.org/10.21273/JASHS.130.6.806
- Nieto-Serrano, E., Campillo, C., Prieto, M.H., Blanco, V., 2026. Impact of altitude and irrigation on sweet cherry tree water status and fruit production in the mountain agroecosystem of the Jerte valley. Scientia Horticulturae, 359, 114765. https://doi.org/10.1016/j.scienta.2026.114765
- Osterman, J., Benton, F., Hellström, S., Luderer-Pflimpfl, M., Pöpel-Eisenbrandt, A.-K., Stoykova Wild, B., Theodorou, P., Ulbricht, C., Paxton, R.J., 2023. Mason bees and honey bees synergistically enhance fruit set in sweet cherry orchards. Ecology and Evolution, 13, e10289. https://doi.org/10.1002/ece3.10289
- Palma, M., Sepúlveda, Á., Yuri, J.A., 2023. Effect of plastic roof and high tunnel on microclimate, physiology, vegetative growth and fruit characteristics of 'Santina' sweet cherry. Scientia Horticulturae, 317, 112037. https://doi.org/10.1016/j.scienta.2023.112037
- Parveze, M.U., Mir, M.M., Rehman, M.U., Iqbal, U., Khan, S.Q., Khan, F.A., Khan, I., Qayoom, S., Mushtaq, I., Shah, H.K., Gaafar, A.-R.Z., Kaushik, P., 2024. Regulation of crop load and quality in sweet cherry cv. 'Sweet Heart' using blossom thinning. Folia Horticulturae, 36(2), 1-11. https://doi.org/10.2478/fhort-2024-0020
- Penzel, M., Möhler, M., Weltzien, C., Herppich, W.B., Zude-Sasse, M., 2020. Estimation of daily carbon demand in sweet cherry (Prunus avium L.) production. Journal of Applied Botany and Food Quality, 93, 149-158. https://doi.org/10.5073/JABFQ.2020.093.019
- Qiu, Z., Wen, Z., Hou, Q., Qiao, G., Yang, K., Hong, Y., Wen, X., 2021. Cross-talk between transcriptome, phytohormone and HD-ZIP gene family analysis illuminates the molecular mechanism underlying fruitlet abscission in sweet cherry (Prunus avium L.). BMC Plant Biology, 21, 173. https://doi.org/10.1186/s12870-021-02940-8
- Radicevic, S., Maric, S., Cerovic, R., Djordjevic, M., 2013. Assessment of self-(in) compatibility in some sweet cherry (Prunus avium L.) genotypes. Genetika, 45(3), 939-952. https://doi.org/10.2298/GENSR1303939R
- Ranjan, R., Sinha, R., Khot, L.R., Whiting, M., 2022. Thermal-RGB imagery and in-field weather sensing derived sweet cherry wetness prediction model. Scientia Horticulturae, 294, 110782. https://doi.org/10.1016/j.scienta.2021.110782
- Rojas, G., Fernández, E., Whitney, C., Luedeling, E., Cuneo, I.F., 2021. Adapting sweet cherry orchards to extreme weather events—Decision analysis in support of farmers’ investments in Central Chile. Agricultural Systems, 187, 103031. https://doi.org/10.1016/j.agsy.2020.103031
- Ruiz-Aracil, M.C., Valverde, J.M., Beltrà, A., Lorente-Mento, J.M., Carrión-Antolí, A., Valero, D., Guillén, F., 2024. Enhancing sweet cherry resilience to spring frost and rain-induced cracking with pre-harvest melatonin treatments. Current Plant Biology, 40, 100388. https://doi.org/10.1016/j.cpb.2024.100388
- Salazar-Gutiérrez, M.R., Chaves, B., Anothai, J., Whiting, M., Hoogenboom, G., 2014. Variation in cold hardiness of sweet cherry flower buds through different phenological stages. Scientia Horticulturae, 172, 161-167. https://doi.org/10.1016/j.scienta.2014.04.002
- Santolaria, N., Castel, L., Rodrigo, J., Fadón, E., 2026. Warming winters and cultivar resilience in sweet cherry: Agroclimatic requirements and future suitability under Mediterranean-continental conditions. Agricultural and Forest Meteorology, 383, 111138. https://doi.org/10.1016/j.agrformet.2026.111138
- Schmitz-Eiberger, M.A., Blanke, M.M., 2012. Bioactive components in forced sweet cherry fruit (Prunus avium L.), antioxidative capacity and allergenic potential as dependent on cultivation under cover. LWT - Food Science and Technology, 46(2), 388-392. https://doi.org/10.1016/j.lwt.2011.12.015
- Schumann, C., Winkler, A., Knoche, M., 2022. Calcium decreases cell wall swelling in sweet cherry fruit. Scientific Reports, 12, 16496. https://doi.org/10.1038/s41598-022-20266-9
- Schuster, M., 2012. Incompatible (S-) genotypes of sweet cherry cultivars (Prunus avium L.). Scientia Horticulturae, 148, 59-73. https://doi.org/10.1016/j.scienta.2012.09.012
- Siopa, C., Castro, S., Loureiro, J., Gaspar, H., Lopes, S., 2026. Flowering synchrony and cultivar compatibility as key factors for optimising sweet cherry pollination and production. Annals of Applied Biology. https://doi.org/10.1111/aab.70044
- Sønsteby, A., Heide, O.M., 2019. Temperature effects on growth and floral initiation in sweet cherry (Prunus avium L.). Scientia Horticulturae, 257, 108762. https://doi.org/10.1016/j.scienta.2019.108762
- Varaldo, A., Giacalone, G., 2025a. Enhancing cracking resistance and post-harvest quality of sweet cherries (Prunus avium L.) through calcium and potassium-based foliar treatments. Horticulturae, 11(1), 30. https://doi.org/10.3390/horticulturae11010030
- Vignati, E., Caccamo, M., Dunwell, J.M., Simkin, A.J., 2024. Morphological changes to fruit development induced by GA3 application in sweet cherry (Prunus avium L.). Plants, 13(15), 2052. https://doi.org/10.3390/plants13152052
- Villar, L., Lienqueo, I., Llanes, A., Rojas, P., Pérez, J., Correa, F., Sagredo, B., Masciarelli, O., Luna, V., Almada, R., Silva, H., 2020. Comparative transcriptomic analysis reveals novel roles of transcription factors and hormones during the flowering induction and floral bud differentiation in sweet cherry trees (Prunus avium L. cv. Bing). PLOS ONE, 15(3), e0230110. https://doi.org/10.1371/journal.pone.0230110
- Vimont, N., Fouché, M., Campoy, J.A., Tong, M., Arkoun, M., Yvin, J.-C., Wigge, P.A., Dirlewanger, E., Cortijo, S., Wenden, B., 2019. From bud formation to flowering: Transcriptomic state defines the cherry developmental phases of sweet cherry bud dormancy. BMC Genomics, 20, 974. https://doi.org/10.1186/s12864-019-6348-z
- Wang, L., Sun, W., Liu, X., Xu, Y., Lyu, Z., Liu, R., Jiu, S., Zhang, C., Wang, J., 2023. Genome-wide identification of the NCED gene family and functional characterization of PavNCED5 related to bud dormancy in sweet cherry. Scientia Horticulturae, 319, 112186. https://doi.org/10.1016/j.scienta.2023.112186
- Wani, A.A., Singh, P., Gul, K., Wani, M.H., Langowski, H.-C., 2014. Sweet cherry (Prunus avium): Critical factors affecting the composition and shelf life. Food Packaging and Shelf Life, 1(1), 86-99. https://doi.org/10.1016/j.fpsl.2014.01.005
- Whiting, M.D., Ophardt, D., 2005. Comparing novel sweet cherry crop load management strategies. HortScience, 40(5), 1271-1275.
- Winkler, A., Blumenberg, I., Schürmann, L., Knoche, M., 2020. Rain cracking in sweet cherries is caused by surface wetness, not by water uptake. Scientia Horticulturae, 269, 109400. https://doi.org/10.1016/j.scienta.2020.109400
- Wójcik, P., Morgaś, H., 2013. Response of 'Burlat' sweet cherry trees to postharvest sprays of nitrogen, boron and zinc. Journal of Plant Nutrition, 36(3), 503-514. https://doi.org/10.1080/01904167.2012.748071
- Xu, H., Ediger, D., Sharifi, M., 2023. Horticultural practices in early spring to mitigate the adverse effect of low temperature on fruit set in ‘Lapins’ sweet cherry. Plants, 12(3), 468. https://doi.org/10.3390/plants12030468
- Zhang, L., Ferguson, L., Whiting, M.D., 2018. Temperature effects on pistil viability and fruit set in sweet cherry. Scientia Horticulturae, 241, 8-17. https://doi.org/10.1016/j.scienta.2018.06.039
Fonte immagine apertura: Stefano Lugli