



Preharvest factors for optimal sweet cherry production
Fruit development: water, crop load, canopy, and nutrition
Sweet cherry fruit growth is commonly described as a double-sigmoid pattern organized into three physiological phases.
Phase I is dominated by cell division and initial mesocarp growth; during Phase II, external growth slows while embryo development and endocarp hardening proceed; and in Phase III, or the fruit-enlargement phase, intense cell expansion resumes, associated with water influx, sugar accumulation, color development, and commercial ripening.
This sequence explains why irrigation, crop load, canopy light environment, and nutrition do not exert the same effect across phases: an intervention that is beneficial in one window may be irrelevant or even counterproductive in another (Wani et al., 2014; Vignati et al., 2024).
During the enlargement phase, fruit vascular dynamics undergo a critical shift.
Water management
Xylem water flow progressively decreases until it becomes almost negligible at maturity, whereas phloem contribution increases and becomes the dominant pathway for water and assimilate entry.
This transition reduces the capacity of mature fruit to buffer abrupt changes in water status and helps explain its high sensitivity to water fluctuations during the final stages of development (Brüggenwirth et al., 2016).
Water management during ripening must be interpreted from this physiology.
Severe water deficit can limit growth, reduce fruit size, and compromise commercial quality; conversely, excessive water availability or abrupt alternation between deficit and rehydration can create conditions that predispose fruit to microcracking and cracking.
Irrigation scheduling
In mountain agroecosystems, elevation and irrigation scheduling condition tree water status, phenology, and fruit size, reinforcing the need to adjust water management to cultivar, rootstock, soil, and local atmospheric demand (Nieto-Serrano et al., 2026).
In addition, vascular contribution to water supply may participate in certain cracking types, especially when mature fruit, high varietal susceptibility, and rainfall or rehydration events coincide (Measham et al., 2010; Knoche et al., 2025).
Irrigation scheduling should therefore be linked to phenological stage, soil water storage, evaporative demand, tree water status, and rainfall forecast.
In practice, the highest risk is often not a single level of water availability, but a sequence of stress and rapid rehydration during the late enlargement and ripening period.
Crop load
Monitoring should aim to reduce abrupt physiological oscillations while maintaining sufficient water supply for fruit growth and canopy function.
Crop load and canopy architecture condition source-sink relationships and fruit microclimate.
High crop load increases competition among fruit, can reduce fruit size, and can modify soluble-solids accumulation; excessively low crop load, however, can favor vegetative vigor, shading, and imbalances in resource distribution.
Pruning, canopy design, and crop-load regulation must be calibrated according to cultivar, rootstock, vigor, water availability, and commercial target, without extrapolating responses obtained in one training system, elevation, or protection infrastructure to all orchards.
Source-sink balance
The relevant target is not crop reduction per se, but a functional balance between fruit number, leaf area, light interception, and assimilate supply (Whiting & Ophardt, 2005; Wani et al., 2014; Blanco et al., 2019; Penzel et al., 2020; Nieto-Serrano et al., 2026).
This source-sink interpretation has direct management consequences.
Estimated daily leaf-area demand per fruit increases during development and can reach high values during Stage III, with cultivar-specific estimates reported for 'Bellise' and 'Regina'.
Crop-load regulation, blossom thinning, and fruit-load adjustment can improve fruit size and selected quality attributes in some systems, yet responses are not universal and may vary with cultivar, rootstock, fruiting density, canopy capacity, cover system, and market price structure.
Mineral nutrition
Crop-load decisions should therefore be made against a defined commercial target rather than as a generic thinning prescription (Whiting & Ophardt, 2005; Einhorn et al., 2011; Penzel et al., 2020; González-Villagra et al., 2024; Parveze et al., 2024).
Mineral nutrition should be interpreted functionally by phenological stage.
Potassium participates in sugar transport, osmotic regulation, and quality during fruit enlargement, making its availability particularly relevant during Phase III.
Calcium contributes to cell-wall integrity and to reduced epidermal cell-wall swelling, although its low mobility toward the fruit and wash-off by rain limit the effectiveness of late applications.
Nutrient response
Responses to foliar treatments must therefore be interpreted according to formulation, timing, coverage, cultivar, cuticle, environmental conditions, and actual wetting risk (Wani et al., 2014; Schumann et al., 2022; Kafle et al., 2016; Varaldo & Giacalone, 2025a).
Table 5 summarizes the nutrients most relevant to fruit development and quality.
The relationship among nutrition, firmness, and cracking must be formulated cautiously.
Foliar calcium and potassium treatments can improve mechanical properties and selected quality attributes under specific experimental conditions, but they should not be presented as a universal solution to cracking.
Cracking nutrition
Their effect depends on cultivar, maturity stage, cuticular susceptibility, previous microcracking, water management, formulation, rain wash-off, and actual wetting pressure.
In this section, nutrition is therefore addressed as functional support for growth and quality; the specific physiology of rain-induced cracking is developed in Section 8 (Schumann et al., 2022; Kafle et al., 2016; Knoche et al., 2025; Varaldo & Giacalone, 2025a).
Table 5. Key nutrients by phenological stage and production function
| Nutrient | Critical window | Main function | Management risk | Key sources |
|---|---|---|---|---|
| Nitrogen (N) | Postharvest, budbreak, and early growth. | Leaf area, reserves, and photosynthetic capacity. | Excess: vigor, shading, softer fruit, and lower floral differentiation. | Michailidis et al. (2018); Wani et al. (2014) |
| Boron (B) | Prebloom, bloom, and fruit set. | Pollen, pollen tube, and cell wall. | Narrow margin between deficiency and toxicity; diagnosis required. | Hedhly et al. (2004); Xu et al. (2023) |
| Zinc (Zn) | Bud swelling and budbreak. | Early growth, enzyme activity, and bud fertility. | Applications require local validation. | Michailidis et al. (2018); Wani et al. (2014) |
| Potassium (K) | Fruit enlargement and ripening. | Sugar transport, osmotic regulation, and quality. | Imbalance with Ca/Mg or increased turgor under poor water management. | Wani et al. (2014); Varaldo & Giacalone (2025a) |
| Calcium (Ca) | Fruit set, cell division, and ripening. | Cell wall, firmness, and lower epidermal swelling. | Low mobility, rain wash-off, and variable foliar response; not a stand-alone solution. | Schumann et al. (2022); Kafle et al. (2016); Varaldo & Giacalone (2025a) |
Note: Nutrient management should be supported by leaf and soil analysis, local diagnosis, cultivar, rootstock, and current regulations.
Ripening, rainfall, cracking, and protected production systems
Ripening concentrates commercial value and vulnerability.
In this final phase, the fruit expresses color, size, firmness, soluble solids, acidity, and flavor, but it also becomes more sensitive to rain, hail, prolonged surface wetness, cracking, microcracking, and associated rots.
Cracking incidence and type vary according to genotype, season, maturity stage, and environmental conditions during the rainfall event, explaining why the same precipitation event can generate very different responses among cultivars, plots, or years (Measham et al., 2009; Blanco et al., 2019; Rojas et al., 2021).
Rain-induced cracking should not be interpreted as the simple consequence of massive water uptake and uniform increases in fruit turgor.
Surface wetness
Available experimental evidence indicates that localized surface wetness is a critical trigger: water retained on the skin preferentially penetrates through microcracks and cuticular discontinuities, promoting localized damage in epidermal and hypodermal tissues that can propagate into visible cracks (Winkler et al., 2020; Knoche et al., 2022).
Vascular contribution can be relevant for specific crack types, particularly side cracking, but it should be formulated as a complementary, context-dependent pathway rather than as the sole explanation of the phenomenon (Measham et al., 2010).
Cracking susceptibility integrates genetic properties, maturity stage, fruit size and firmness, cuticle characteristics, previous microcracking, tissue calcium, and surface wetness duration.
This integration connects the variability observed among genotypes and seasons with local mechanisms of skin-integrity loss.
Zipper model
In full-sib families contrasting for cracking susceptibility, tolerance has been associated with cuticular traits, microcracking, and calcium, reinforcing the need to interpret cracking as a skin-structure problem rather than merely as a general fruit water response (Measham et al., 2009; Knoche et al., 2025).
The “zipper” model provides a mechanistic framework for explaining damage propagation.
According to this model, surface wetness and microcracking facilitate localized water entry, lysis of epidermal or subepidermal cells, and release of organic acids, especially malate, into the apoplast.
This process can extract calcium from neighboring cell walls, increase wall swelling, reduce cell adhesion, and promote progressive unzipping of the skin.
Mechanistic sequence
The model should not be treated as a closed explanation for all cultivars and conditions, but as an integrative hypothesis linking cuticle, surface water, calcium, cell wall, and crack propagation (Knoche & Winkler, 2019; Schumann et al., 2022; Knoche et al., 2022).
Figure 4 synthesizes this mechanistic sequence and its applied interpretation.
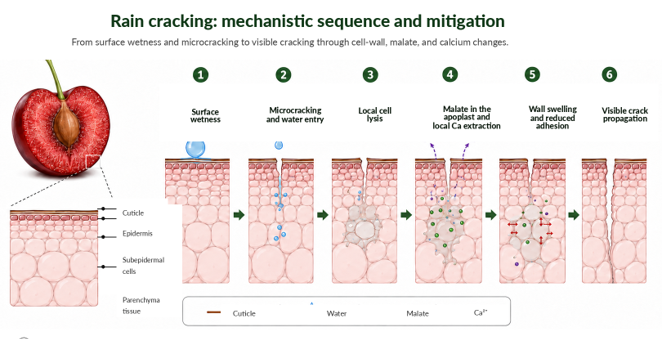
Figure 4. Proposed mechanistic sequence for rain-induced cracking in sweet cherry.
The scheme synthesizes the transition from surface wetness and cuticular microcracking to localized water entry, cell lysis, malate release into the apoplast, local calcium extraction, wall swelling, loss of cell adhesion, and visible crack propagation.
Mitigation strategies
Mitigation strategies must act on several points of the system: reducing fruit-surface wetness duration, avoiding abrupt water-status oscillations, maintaining a ventilated canopy, decreasing direct exposure to rain or hail, and applying nutrition programs or preharvest treatments only when diagnosis and local evidence support them.
Foliar calcium or potassium applications can improve mechanical properties and reduce cracking susceptibility in specific trials, but their effectiveness depends on formulation, dose, timing, coverage, rain wash-off, cultivar, and maturity stage (Kafle et al., 2016; Schumann et al., 2022; Varaldo & Giacalone, 2025a).
Similarly, treatments with abscisic acid, methyl jasmonate, or melatonin have shown favorable effects in specific contexts, although they should be considered strategies conditioned by cultivar, phenological stage, dose, regulatory status, and actual rainfall pressure (Balbontín et al., 2018; Ruiz-Aracil et al., 2024).
Because surface wetness is central to cracking risk, monitoring technologies can add value when they estimate fruit wetness rather than only rainfall.
Protected systems
Thermal-RGB imagery combined with in-field weather sensing has produced cultivar-specific wetness prediction models, illustrating how fruit-surface temperature and local microclimate can be integrated into decision support for drying, spraying, or rain-protection strategies (Ranjan et al., 2022).
Plastic covers, high tunnels, and other protected systems reduce direct contact between rainfall and fruit and can decrease losses from cracking or hail.
However, they are not neutral structures: they modify radiation, temperature, relative humidity, ventilation, vigor, ripening, quality, and disease management.
High tunnels have been shown to protect against rainfall and modify physiological and production variables, whereas forcing systems under cover can alter radiation, temperature, firmness, soluble solids, and bioactive compounds.
Production system
Recent work under Mediterranean conditions has also shown that roof and high-tunnel systems can reduce incident radiation, modify canopy-level microclimate and tree water status, advance harvest, and affect color, firmness, acidity, phenolics, and antioxidant capacity.
Physical protection should therefore be evaluated as a modification of the production system and as a risk-management investment, not merely as a barrier against an isolated weather event (Schmitz-Eiberger & Blanke, 2012; Blanco et al., 2019; Rojas et al., 2021; Palma et al., 2023).
Climate change and adaptation of the production system
Climate change modifies the agroclimatic framework in which sweet cherry production is built because it simultaneously affects winter chill accumulation, spring heat, phenological synchrony, and the frequency of extreme events during bloom, fruit set, and ripening.
In sweet cherry, warming does not necessarily imply a simple phenological advance.
Dormancy risk
When winter chill is insufficient, endodormancy release can be incomplete, resulting in irregular budbreak, uneven bloom, and lower production stability.
Conversely, when chilling requirements are satisfied, a warmer spring can accelerate heat accumulation and advance bloom, increasing exposure of reproductive organs to late frost.
Adaptation therefore cannot be based on a single thermal indicator; it must consider the interaction among chill, forcing, cultivar, bloom date, and local phenological risk (Fadón et al., 2021; Fadón et al., 2023).
Climate vulnerability is strongly cultivar-specific.
Cultivar viability
Studies of agroclimatic requirements in Spanish and commercial cultivars show that genotypes differ in chill and heat needs and in their capacity to maintain regular bloom under warming scenarios.
Projections under Mediterranean-continental conditions in the Ebro Valley indicate that, by 2050, most evaluated cultivars may still maintain reasonable climatic viability, although cultivars with higher chilling requirements show earlier risks of maladaptation.
Toward the end of the century, especially under high-emission scenarios, even lower-chill cultivars may be at risk of failing to satisfy their chilling needs in some climate models.
These results should not be extrapolated without regional calibration, but they emphasize that cultivar selection and agroclimatic zoning will become increasingly critical decisions (Fadón et al., 2023; Santolaria et al., 2026).
Structural adaptation
Structural adaptation begins before planting.
It includes site, elevation, aspect, cultivar, rootstock, water availability, canopy design, and protection infrastructure.
In mountain agroecosystems such as the Jerte Valley, physiological and meteorological measurements show that higher elevation can reduce evaporative demand, improve tree water status, delay phenology, and extend the fruit-development period, favoring larger commercial fruit sizes compared with lower areas exposed to stronger combined heat and water stress.
This evidence supports the value of elevation as a local adaptation factor, although its use must be interpreted within each production system and its limitations in soil, slope, water, accessibility, and frost risk (Nieto-Serrano et al., 2026).
Protection infrastructure
Protection infrastructure is part of structural adaptation, but it should not be considered neutral.
High tunnels and plastic covers can reduce losses from rain or hail and modify maturity date, but they also alter radiation, temperature, relative humidity, ventilation, vigor, quality, and disease management.
Their usefulness depends on climate risk, cultivar, training system, installation costs, management logistics, and market target.
Investment decisions in covers or other protection technologies should therefore be evaluated as risk-management problems, not merely as one-off responses to a weather event (Blanco et al., 2019; Rojas et al., 2021).
Operational measures
Alongside structural measures, adaptation requires operational measures adjusted each season.
These include hourly monitoring of chill and heat, phenological tracking, irrigation management, crop-load adjustment, pruning to modulate canopy microclimate, and preparedness for frost, rain, or hail.
Irrigation, crop load, and canopy management can help modulate tree water status, fruit microclimate, and exposure to final ripening risks, whereas regulated deficit irrigation and water management must always be formulated according to cultivar, rootstock, soil, phenological phase, and water availability.
Operational adaptation is therefore not the application of fixed recipes, but the reduction of physiological oscillations that increase fruit and tree vulnerability (Blanco et al., 2019; Nieto-Serrano et al., 2026).
Decision support
Within this framework, instrumental monitoring and decision-support systems can help translate physiological understanding into more precise operational decisions.
Integration of hourly data on chill, heat, rainfall, humidity, wet-bulb temperature, fruit-surface wetness, phenological monitoring, tree water-status indicators, and probabilistic risk assessment can guide irrigation, frost protection, anticipation of surface wetness associated with cracking, and investment in covers or nets under local conditions.
These tools do not replace agronomic judgment or eliminate uncertainty, but they improve decision traceability when interpreted together with cultivar, rootstock, microclimate, fruit maturity, available infrastructure, and commercial target (Fadón et al., 2021; Fadón et al., 2023; Rojas et al., 2021; Ranjan et al., 2022; Nieto-Serrano et al., 2026).
Overall, adaptation of sweet cherry to climate change requires integration of thermal models, tree physiology, economic risk, and localized management.
Adaptation strategy
The most robust strategies will combine agroclimatic cultivar zoning, selection of adapted cultivars and rootstocks, physical protection when justified, efficient water management, phenological monitoring, and economic decision analysis.
This interpretation closes the chapter with a central idea: optimal sweet cherry production will depend not only on increasing yield or quality under average conditions, but on designing systems capable of maintaining production stability under more irregular winters, more uncertain springs, and more frequent extreme events (Rojas et al., 2021; Fadón et al., 2023; Santolaria et al., 2026).
Table 6 summarizes the main preharvest factors, their critical timing, expected production effect, and associated technical orientation.
Its interpretation must remain contextual and traceable to the available evidence.
Preharvest matrix
Table 6. Synthetic matrix of preharvest factors and contextualized management orientation
| Timing | Critical factor | Productive effect | Technical orientation |
|---|---|---|---|
| Establishment | Site, elevation, aspect, drainage, cultivar, rootstock, and infrastructure. | Defines chill, frost, vigor, water, root health, and exposure to extreme events. | Prioritize local structural adaptation and do not separate cultivar from rootstock, microclimate, and water availability. |
| Previous cycle | Summer heat, reserves, vigor, and nutrition. | Conditions floral induction and bud quality. | Avoid severe stress and excessive vigor; maintain an illuminated canopy. |
| Dormancy-budbreak | Chill CP/CH/CU and forcing GDH. | Regulates bloom uniformity and date. | Use hourly data and explicit models; avoid universal thresholds. |
| Bloom-fruit set | Frost, compatibility, EPP, and pollinators. | Determines fertilization, fruit set, and initial crop load. | Design compatible pollinizers and activate mitigation based on wet-bulb temperature and phenophase. |
| Enlargement-ripening | Water, evaporative demand, crop load, K-Ca balance, rainfall, cover, and microclimate. | Defines size, quality, firmness, cracking, and final losses. | Adjust irrigation, crop load, canopy, and nutrition; evaluate physical protection according to risk and cost. |
| Climate adaptation | Chill, forcing, elevation, water, frost, rain, and hail. | Conditions cultivar viability, phenological stability, and economic risk. | Combine zoning, hourly monitoring, operational management, and decision analysis. |
Note: This matrix is intended as a contextual decision framework and should not be used as a fixed prescription.
Operationally, the matrix is best used as a seasonal monitoring loop: first define the structural risk of the site and cultivar, then update chill and forcing accumulation during dormancy, intensify frost and pollination decisions during bloom, adjust water and crop load during fruit development, and reassess rain, wetness, cracking, and harvest risk during ripening.
This sequence turns the physiological chain described in the chapter into a traceable management process.
Conclusions
Optimal sweet cherry production should be understood as a continuous physiological and agronomic chain in which the commercial yield of a season depends on processes initiated during the previous cycle.
Floral induction, bud quality, winter chill accumulation, spring heat, frost sensitivity, bloom, pollination, and fruit set operate as interdependent stages.
Commercial yield
Productive success therefore cannot be attributed to a single management practice or to a single thermal indicator, but to coherence among cultivar, rootstock, site, phenology, microclimate, and agronomic decisions adjusted to each system.
During fruit development, management of water, crop load, canopy, and mineral nutrition conditions the final expression of fruit size, firmness, color, soluble solids, and commercial quality.
Ripening, however, concentrates both the highest value and the highest vulnerability of the crop.
At this stage, reduced xylem contribution, phloem-dominated inflow, solute accumulation, cuticular tension, and microcracking increase fruit sensitivity to rain, surface wetness, cracking, hail, and rots.
Final risks
Quality built over months can be lost rapidly if final ripening risks are not anticipated through water management, physical protection, canopy ventilation, contextualized nutrition, and adjusted harvest decisions.
Under climate change, the production stability of sweet cherry will increasingly depend on the capacity to adapt orchard systems to more irregular winters, more uncertain springs, greater combined heat and water stress, and more frequent extreme events.
This adaptation should be organized at two complementary levels: structural measures, such as selection of region, elevation, cultivar, rootstock, planting design, and protection infrastructure; and operational measures, such as phenological monitoring, irrigation adjustment, crop-load regulation, canopy management, and response to frost, rain, or hail.
None of these measures should be interpreted as a universal recipe, but as part of a local strategy based on diagnosis, evidence, and risk analysis.
Systems approach
Overall, preharvest management of sweet cherry should move from isolated interventions toward a systems approach.
Stable commercial production is achieved not only by maximizing yield or quality under average conditions, but by reducing tree and fruit vulnerability during the most sensitive phenological windows.
Integrating physiology, agroclimatic data, cultivar-specific thresholds, local monitoring, and decision analysis will be essential for converting the biological potential of sweet cherry into high-quality fruit, resilient production, and sustainable commercial yield.
Jesús Alonso¹,²
¹ Institute of Food Science, Technology and Nutrition (ICTAN-CSIC), Spanish National Research Council (CSIC), c/ José Antonio Novais, 6, 28040 Madrid, Spain.
² University Research Institute for Agricultural Resources (INURA), Universidad de Extremadura, Edificio de los Institutos Universitarios de Investigación, Avenida de la Investigación s/n, 06006 Badajoz, Spain.
References
- Alburquerque, N., García-Montiel, F., Carrillo, A., Burgos, L., 2008. Chilling and heat requirements of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements. Environmental and Experimental Botany, 64(2), 162-170. https://doi.org/10.1016/j.envexpbot.2008.01.003
- Ayala, M., Lang, G.A., 2015. 13C-photoassimilate partitioning in sweet cherry (Prunus avium) during early spring. Ciencia e Investigación Agraria, 42(2), 191-203. https://doi.org/10.4067/S0718-16202015000200006
- Azizi Gannouni, T., Campoy, J.A., Quero-García, J., Barreneche, T., Arif, A., Albouchi, A., Ammari, Y., 2017. Dormancy related traits and adaptation of sweet cherry in Northern Africa: A case of study in two Tunisian areas. Scientia Horticulturae, 219, 272-279. https://doi.org/10.1016/j.scienta.2017.03.013
- Balbontín, C., Gutiérrez, C., Wolff, M., Figueroa, C.R., 2018. Effect of abscisic acid and methyl jasmonate preharvest applications on fruit quality and cracking tolerance of sweet cherry. Chilean Journal of Agricultural Research, 78(3), 438-446. https://doi.org/10.4067/S0718-58392018000300438
- Blanco, V., Zoffoli, J.P., Ayala, M., 2019. High tunnel cultivation of sweet cherry (Prunus avium L.): Physiological and production variables. Scientia Horticulturae, 251, 108-117. https://doi.org/10.1016/j.scienta.2019.02.023
- Bonomelli, C., Bonilla, C., Acuña, E., Artacho, P., 2012. Seasonal pattern of root growth in relation to shoot phenology and soil temperature in sweet cherry trees (Prunus avium): A preliminary study in central Chile. Ciencia e Investigación Agraria, 39(1), 127-136.
- Brüggenwirth, M., Winkler, A., Knoche, M., 2016. Xylem, phloem, and transpiration flows in developing sweet cherry fruit. Trees, 30, 1821-1830. https://doi.org/10.1007/s00468-016-1415-4
- Choi, C., Tao, R., Andersen, R.L., 2002. Identification of self-incompatibility alleles and pollen incompatibility groups in sweet cherry by PCR-based S-allele typing and controlled pollination. Euphytica, 123, 9-20. https://doi.org/10.1023/A:1014403802677
- Djordjevic, B.S., Djurovic, D.B., Zec, G.N., Boskov, D.D., 2021. The influence of rootstocks on the sensitivity of flower buds to frost and the main properties of the 'Carmen' sweet cherry cultivar. Journal of Agricultural Sciences (Belgrade), 66(4), 351-358. https://doi.org/10.2298/JAS2104351D
- Dziedzic, E., Bieniasz, M., Kowalczyk, B., 2019. Morphological and physiological features of sweet cherry floral organ affecting the potential fruit crop in relation to the rootstock. Scientia Horticulturae, 251, 127-135. https://doi.org/10.1016/j.scienta.2019.03.013
- Eeraerts, M., Meeus, I., Van Den Berge, S., Smagghe, G., 2017. Landscapes with high intensive fruit cultivation reduce wild pollinator services to sweet cherry. Agriculture, Ecosystems & Environment, 239, 342-348. https://doi.org/10.1016/j.agee.2017.01.031
- Eeraerts, M., Smagghe, G., Meeus, I., 2020a. Bumble bee abundance and richness improves honey bee pollination behaviour in sweet cherry. Basic and Applied Ecology, 43, 27-33. https://doi.org/10.1016/j.baae.2019.11.004
- Eeraerts, M., Vanderhaegen, R., Smagghe, G., Meeus, I., 2020b. Pollination efficiency and foraging behaviour of honey bees and non-Apis bees to sweet cherry. Agricultural and Forest Entomology, 22, 75-82. https://doi.org/10.1111/afe.12363
- Einhorn, T.C., Laraway, D., Turner, J., 2011. Crop load management does not consistently improve crop value of 'Sweetheart'/'Mazzard' sweet cherry trees. HortTechnology, 21(5), 546-553.
- Fadón, E., Fernández, E., Luedeling, E., Rodrigo, J., 2023. Agroclimatic requirements and adaptation potential to global warming of Spanish cultivars of sweet cherry (Prunus avium L.). European Journal of Agronomy, 145, 126774. https://doi.org/10.1016/j.eja.2023.126774
- Fadón, E., Herrero, M., Rodrigo, J., 2015. Flower development in sweet cherry framed in the BBCH scale. Scientia Horticulturae, 192, 141-147. https://doi.org/10.1016/j.scienta.2015.05.027
- Fadón, E., Herrero, M., Rodrigo, J., 2018a. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science, 9, 171. https://doi.org/10.3389/fpls.2018.00171
- Fadón, E., Rodrigo, J., Herrero, M., 2018b. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees, 32(6), 1583-1594. https://doi.org/10.1007/s00468-018-1735-7
- Fadón, E., Rodrigo, J., Luedeling, E., 2021. Cultivar-specific responses of sweet cherry flowering to rising temperatures during dormancy. Agricultural and Forest Meteorology, 307, 108486. https://doi.org/10.1016/j.agrformet.2021.108486
- Feldmane, D., Sebre, G., Rubauskis, E., 2025. Cultivar and rootstock effect on sweet cherry fruit density and fruit drop. Rural Sustainability Research, 54(349), 79-86. https://doi.org/10.2478/plua-2025-0019
- García, C.B., Díaz-Siefer, P., Smith-Ramírez, C., Montero-Silva, F., Martínez-Harms, J., Murúa, M., Celis-Diez, J.L., 2025. Synergistic effect of honeybees and wild floral visitors in promoting sweet cherry fruit set in central Chile. Biological Research, 58(1), 39. https://doi.org/10.1186/s40659-025-00617-2
- González-Villagra, J., Palacios-Peralta, C., Muñoz-Alarcón, A., Reyes-Díaz, M., Osorio, P., Ribera-Fonseca, A., 2024. Influence of fruit load regulation on harvest and postharvest fruit quality and antioxidant-related parameters in sweet cherry (Prunus avium L.) cv. Regina cultivated under plastic covers in Southern Chile. Plants, 13, 2257. https://doi.org/10.3390/plants13162257
- Götz, K.-P., Naher, J., Fettke, J., Chmielewski, F.-M., 2018. Changes of proteins during dormancy and bud development of sweet cherry (Prunus avium L.). Scientia Horticulturae, 239, 41-49. https://doi.org/10.1016/j.scienta.2018.05.016
- Hedhly, A., Hormaza, J.I., Herrero, M., 2004. Effect of temperature on pollen tube kinetics and dynamics in sweet cherry, Prunus avium (Rosaceae). American Journal of Botany, 91(4), 558-564. https://doi.org/10.3732/ajb.91.4.558
- Hedhly, A., Hormaza, J.I., Herrero, M., 2007. Warm temperatures at bloom reduce fruit set in sweet cherry. Journal of Applied Botany and Food Quality, 81, 158-164. https://doi.org/10.5073/JABFQ.2007.081.025
- Hedhly, A., Hormaza, J.I., Herrero, M., 2009. Flower emasculation accelerates ovule degeneration and reduces fruit set in sweet cherry. Scientia Horticulturae, 119(4), 455-457. https://doi.org/10.1016/j.scienta.2008.08.020
- Holzschuh, A., Dudenhöffer, J.-H., Tscharntke, T., 2012. Landscapes with wild bee habitats enhance pollination, fruit set and yield of sweet cherry. Biological Conservation, 153, 101-107. https://doi.org/10.1016/j.biocon.2012.04.032
- Houghton, E., Watanabe, Y., Neilsen, D., Nelson, L.M., Hannam, K., 2024. Investigating properties of sweet cherry (Prunus avium) flower buds that help promote freezing avoidance by supercooling. Plant Biology, 26(6), 1067-1078. https://doi.org/10.1111/plb.13697
- Kafle, G.K., Khot, L.R., Zhou, J., Bahlol, H.Y., Si, Y., 2016. Towards precision spray applications to prevent rain-induced sweet cherry cracking: Understanding calcium washout due to rain and fruit cracking susceptibility. Scientia Horticulturae, 203, 152-157. https://doi.org/10.1016/j.scienta.2016.03.027
- Karbassioon, A., Yearsley, J., Dirilgen, T., Hodge, S., Stout, J.C., Stanley, D.A., 2023. Responses in honeybee and bumblebee activity to changes in weather conditions. Oecologia, 201, 689-701. https://doi.org/10.1007/s00442-023-05332-x
- Kaufmann, H., Blanke, M.M., 2017. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology, 218, 1-5. https://doi.org/10.1016/j.jplph.2017.07.004
- Kaufmann, H., Blanke, M.M., 2019. Substitution of winter chilling by spring forcing for flowering using sweet cherry as model crop. Scientia Horticulturae, 244, 75-81. https://doi.org/10.1016/j.scienta.2018.09.021
- Kaya, O., Köse, C., 2022. How sensitive are the flower parts of the sweet cherry in sub-zero temperatures? Use of differential thermal analysis and critical temperatures assessment. New Zealand Journal of Crop and Horticultural Science, 50(1), 17-31. https://doi.org/10.1080/01140671.2021.1890156
- Kaya, O., Köse, C., Şahin, M., 2021. The use of differential thermal analysis in determining the critical temperatures of sweet cherry (Prunus avium L.) flower buds at different stages of bud burst. International Journal of Biometeorology, 65(7), 1125-1135. https://doi.org/10.1007/s00484-021-02093-1
- Knoche, M., Grosset-Grange, L., Quero-García, J., Alletru, D., Boutaleb, L., 2025. Cracking susceptibility of full-sibs of a cross of a cracking tolerant and cracking susceptible sweet cherry: Relation to cuticle characteristics, microcracking and calcium. PLOS ONE, 20(1), e0316637. https://doi.org/10.1371/journal.pone.0316637
- Knoche, M., Winkler, A., 2019. The mechanism of rain cracking of sweet cherry fruit. Italus Hortus, 26, 59-65. https://doi.org/10.26353/j.itahort/2019.1.5965
- Knoche, M., Winkler, A., Lang, A., 2022. The unzipping of sweet cherry fruit skin and strategies to prevent it. Italus Hortus, 29(1), 1-13. https://doi.org/10.26353/j.itahort/2022.1.C1
- Laterza, I., Ragone, G., Grossi, G., Cappellari, A., Agostinacchio, M.F., Seclì, G.A., Bari, G., Porro, R., Addante, R., Cornara, D., De Lillo, E., Tamburini, G., 2025. Agricultural intensification at local and landscape scales impacts sweet cherry production through altered pollination services. Agriculture, Ecosystems & Environment, 386, 109586. https://doi.org/10.1016/j.agee.2025.109586
- Mateos-Fierro, Z., Garratt, M.P.D., Fountain, M.T., Ashbrook, K., Westbury, D.B., 2025. Contribution of pollinators to delivering fruit quality in commercial sweet cherry orchards. JSFA Reports. https://doi.org/10.1002/jsf2.70012
- Matzneller, P., Götz, K.-P., Chmielewski, F.-M., 2016. Spring frost vulnerability of sweet cherries under controlled conditions. International Journal of Biometeorology, 60(1), 123-130. https://doi.org/10.1007/s00484-015-1010-1
- McCabe, L.M., Boyle, N.K., Pitts-Singer, T.L., 2024. Osmia lignaria (Hymenoptera: Megachilidae) increase pollination of Washington sweet cherry and pear orchards. Environmental Entomology, 53(4), 698-705. https://doi.org/10.1093/ee/nvae043
- Measham, P.F., Bound, S.A., Gracie, A.J., Wilson, S.J., 2009. Incidence and type of cracking in sweet cherry (Prunus avium L.) are affected by genotype and season. Crop & Pasture Science, 60(10), 1002-1008. https://doi.org/10.1071/CP08410
- Measham, P.F., Gracie, A.J., Wilson, S.J., Bound, S.A., 2010. Vascular flow of water induces side cracking in sweet cherry (Prunus avium L.). Advances in Horticultural Science, 24(4), 243-248. https://www.jstor.org/stable/42883522
- Michailidis, M., Karagiannis, E., Tanou, G., Sarrou, E., Adamakis, I.-D., Karamanoli, K., Martens, S., Molassiotis, A., 2018. Metabolic mechanisms underpinning vegetative bud dormancy release and shoot development in sweet cherry. Environmental and Experimental Botany, 155, 1-11. https://doi.org/10.1016/j.envexpbot.2018.06.024
- Naor, A., Stern, R., Peres, M., Greenblat, Y., Gal, Y., 2005. Timing and severity of postharvest water stress affect following-year productivity and fruit quality of field-grown 'Snow Queen' nectarine. Journal of the American Society for Horticultural Science, 130(6), 806-812. https://doi.org/10.21273/JASHS.130.6.806
- Nieto-Serrano, E., Campillo, C., Prieto, M.H., Blanco, V., 2026. Impact of altitude and irrigation on sweet cherry tree water status and fruit production in the mountain agroecosystem of the Jerte valley. Scientia Horticulturae, 359, 114765. https://doi.org/10.1016/j.scienta.2026.114765
- Osterman, J., Benton, F., Hellström, S., Luderer-Pflimpfl, M., Pöpel-Eisenbrandt, A.-K., Stoykova Wild, B., Theodorou, P., Ulbricht, C., Paxton, R.J., 2023. Mason bees and honey bees synergistically enhance fruit set in sweet cherry orchards. Ecology and Evolution, 13, e10289. https://doi.org/10.1002/ece3.10289
- Palma, M., Sepúlveda, Á., Yuri, J.A., 2023. Effect of plastic roof and high tunnel on microclimate, physiology, vegetative growth and fruit characteristics of 'Santina' sweet cherry. Scientia Horticulturae, 317, 112037. https://doi.org/10.1016/j.scienta.2023.112037
- Parveze, M.U., Mir, M.M., Rehman, M.U., Iqbal, U., Khan, S.Q., Khan, F.A., Khan, I., Qayoom, S., Mushtaq, I., Shah, H.K., Gaafar, A.-R.Z., Kaushik, P., 2024. Regulation of crop load and quality in sweet cherry cv. 'Sweet Heart' using blossom thinning. Folia Horticulturae, 36(2), 1-11. https://doi.org/10.2478/fhort-2024-0020
- Penzel, M., Möhler, M., Weltzien, C., Herppich, W.B., Zude-Sasse, M., 2020. Estimation of daily carbon demand in sweet cherry (Prunus avium L.) production. Journal of Applied Botany and Food Quality, 93, 149-158. https://doi.org/10.5073/JABFQ.2020.093.019
- Qiu, Z., Wen, Z., Hou, Q., Qiao, G., Yang, K., Hong, Y., Wen, X., 2021. Cross-talk between transcriptome, phytohormone and HD-ZIP gene family analysis illuminates the molecular mechanism underlying fruitlet abscission in sweet cherry (Prunus avium L.). BMC Plant Biology, 21, 173. https://doi.org/10.1186/s12870-021-02940-8
- Radicevic, S., Maric, S., Cerovic, R., Djordjevic, M., 2013. Assessment of self-(in) compatibility in some sweet cherry (Prunus avium L.) genotypes. Genetika, 45(3), 939-952. https://doi.org/10.2298/GENSR1303939R
- Ranjan, R., Sinha, R., Khot, L.R., Whiting, M., 2022. Thermal-RGB imagery and in-field weather sensing derived sweet cherry wetness prediction model. Scientia Horticulturae, 294, 110782. https://doi.org/10.1016/j.scienta.2021.110782
- Rojas, G., Fernández, E., Whitney, C., Luedeling, E., Cuneo, I.F., 2021. Adapting sweet cherry orchards to extreme weather events—Decision analysis in support of farmers’ investments in Central Chile. Agricultural Systems, 187, 103031. https://doi.org/10.1016/j.agsy.2020.103031
- Ruiz-Aracil, M.C., Valverde, J.M., Beltrà, A., Lorente-Mento, J.M., Carrión-Antolí, A., Valero, D., Guillén, F., 2024. Enhancing sweet cherry resilience to spring frost and rain-induced cracking with pre-harvest melatonin treatments. Current Plant Biology, 40, 100388. https://doi.org/10.1016/j.cpb.2024.100388
- Salazar-Gutiérrez, M.R., Chaves, B., Anothai, J., Whiting, M., Hoogenboom, G., 2014. Variation in cold hardiness of sweet cherry flower buds through different phenological stages. Scientia Horticulturae, 172, 161-167. https://doi.org/10.1016/j.scienta.2014.04.002
- Santolaria, N., Castel, L., Rodrigo, J., Fadón, E., 2026. Warming winters and cultivar resilience in sweet cherry: Agroclimatic requirements and future suitability under Mediterranean-continental conditions. Agricultural and Forest Meteorology, 383, 111138. https://doi.org/10.1016/j.agrformet.2026.111138
- Schmitz-Eiberger, M.A., Blanke, M.M., 2012. Bioactive components in forced sweet cherry fruit (Prunus avium L.), antioxidative capacity and allergenic potential as dependent on cultivation under cover. LWT - Food Science and Technology, 46(2), 388-392. https://doi.org/10.1016/j.lwt.2011.12.015
- Schumann, C., Winkler, A., Knoche, M., 2022. Calcium decreases cell wall swelling in sweet cherry fruit. Scientific Reports, 12, 16496. https://doi.org/10.1038/s41598-022-20266-9
- Schuster, M., 2012. Incompatible (S-) genotypes of sweet cherry cultivars (Prunus avium L.). Scientia Horticulturae, 148, 59-73. https://doi.org/10.1016/j.scienta.2012.09.012
- Siopa, C., Castro, S., Loureiro, J., Gaspar, H., Lopes, S., 2026. Flowering synchrony and cultivar compatibility as key factors for optimising sweet cherry pollination and production. Annals of Applied Biology. https://doi.org/10.1111/aab.70044
- Sønsteby, A., Heide, O.M., 2019. Temperature effects on growth and floral initiation in sweet cherry (Prunus avium L.). Scientia Horticulturae, 257, 108762. https://doi.org/10.1016/j.scienta.2019.108762
- Varaldo, A., Giacalone, G., 2025a. Enhancing cracking resistance and post-harvest quality of sweet cherries (Prunus avium L.) through calcium and potassium-based foliar treatments. Horticulturae, 11(1), 30. https://doi.org/10.3390/horticulturae11010030
- Vignati, E., Caccamo, M., Dunwell, J.M., Simkin, A.J., 2024. Morphological changes to fruit development induced by GA3 application in sweet cherry (Prunus avium L.). Plants, 13(15), 2052. https://doi.org/10.3390/plants13152052
- Villar, L., Lienqueo, I., Llanes, A., Rojas, P., Pérez, J., Correa, F., Sagredo, B., Masciarelli, O., Luna, V., Almada, R., Silva, H., 2020. Comparative transcriptomic analysis reveals novel roles of transcription factors and hormones during the flowering induction and floral bud differentiation in sweet cherry trees (Prunus avium L. cv. Bing). PLOS ONE, 15(3), e0230110. https://doi.org/10.1371/journal.pone.0230110
- Vimont, N., Fouché, M., Campoy, J.A., Tong, M., Arkoun, M., Yvin, J.-C., Wigge, P.A., Dirlewanger, E., Cortijo, S., Wenden, B., 2019. From bud formation to flowering: Transcriptomic state defines the cherry developmental phases of sweet cherry bud dormancy. BMC Genomics, 20, 974. https://doi.org/10.1186/s12864-019-6348-z
- Wang, L., Sun, W., Liu, X., Xu, Y., Lyu, Z., Liu, R., Jiu, S., Zhang, C., Wang, J., 2023. Genome-wide identification of the NCED gene family and functional characterization of PavNCED5 related to bud dormancy in sweet cherry. Scientia Horticulturae, 319, 112186. https://doi.org/10.1016/j.scienta.2023.112186
- Wani, A.A., Singh, P., Gul, K., Wani, M.H., Langowski, H.-C., 2014. Sweet cherry (Prunus avium): Critical factors affecting the composition and shelf life. Food Packaging and Shelf Life, 1(1), 86-99. https://doi.org/10.1016/j.fpsl.2014.01.005
- Whiting, M.D., Ophardt, D., 2005. Comparing novel sweet cherry crop load management strategies. HortScience, 40(5), 1271-1275.
- Winkler, A., Blumenberg, I., Schürmann, L., Knoche, M., 2020. Rain cracking in sweet cherries is caused by surface wetness, not by water uptake. Scientia Horticulturae, 269, 109400. https://doi.org/10.1016/j.scienta.2020.109400
- Wójcik, P., Morgaś, H., 2013. Response of 'Burlat' sweet cherry trees to postharvest sprays of nitrogen, boron and zinc. Journal of Plant Nutrition, 36(3), 503-514. https://doi.org/10.1080/01904167.2012.748071
- Xu, H., Ediger, D., Sharifi, M., 2023. Horticultural practices in early spring to mitigate the adverse effect of low temperature on fruit set in ‘Lapins’ sweet cherry. Plants, 12(3), 468. https://doi.org/10.3390/plants12030468
- Zhang, L., Ferguson, L., Whiting, M.D., 2018. Temperature effects on pistil viability and fruit set in sweet cherry. Scientia Horticulturae, 241, 8-17. https://doi.org/10.1016/j.scienta.2018.06.039