Il cancro batterico è una delle principali malattie che colpiscono il ciliegio, associata a un complesso di batteri del genere Pseudomonas, essendo Pseudomonas syringae pv. syringae (Pss) il principale agente causa della malattia in Cile. Provoca danni in tutti gli stadi della pianta, causando la morte delle giovani piante in vivaio e una riduzione della resa nelle piante adulte che può raggiungere il 50% della produzione, oltre a una marcata riduzione della vita utile del frutteto.
I fattori coinvolti nello sviluppo della malattia sono molteplici e si intrecciano per innescare l'infezione e favorire il successivo insediamento del complesso batterico in diversi organi della pianta. Tra questi: l'età della pianta, il grado di virulenza dei ceppi batterici, le condizioni ambientali, la presenza di altri agenti infettivi, l'azione di stress abiotici come l'acqua e, infine, la varietà utilizzata.
Se a ciò si aggiunge che oggi il controllo curativo del cancro batterico si basa principalmente su trattamenti ripetuti con prodotti a base di rame e che il batterio è in grado di acquisire ed esprimere meccanismi di resistenza e di generare ceppi sempre più tolleranti al rame, allora è prioritario concentrare il controllo attraverso strategie di prevenzione dell'infezione e di riduzione della diffusione del batterio.
Attualmente, per limitare la diffusione del batterio nel frutteto, si consiglia di rimuovere i rami e i cancri colpiti. La diffusione può avvenire attraverso l'uso di materiale vegetale infetto (ad esempio, tramite innesto), la pioggia, il vento e gli strumenti di manipolazione contaminati (ad esempio, durante la potatura).
La Pss penetra nella pianta attraverso aperture naturali come gli stomi e le lenticelle, o artificiali come le ferite causate dalla caduta delle foglie, le fessure di crescita, le ferite causate dal sistema di conduzione o le microinfezioni generate dalla capacità del batterio di congelare l'acqua (nucleazione del ghiaccio) a temperature appena superiori a 0°C.
Suscettibilità varietale differenziata
Per quanto riguarda l'esportazione di ciliegie, le principali varietà prodotte ed esportate in Cile nella stagione 2022/23 sono state, in ordine di importanza: Lapins, Santina, Regina e Bing (Asoex, 2023). Tuttavia, in base alle osservazioni dei sintomi nei frutteti commerciali e ai test di infezione in laboratorio, Bing è la varietà con la maggiore suscettibilità al cancro batterico, seguita da Regina, Lapins e, infine, Santina.
È importante considerare che il grado di suscettibilità differenziata all'afta batterica presente nelle diverse varietà di ciliegio è principalmente legato alla capacità della pianta di rilevare rapidamente la presenza del patogeno nei tessuti interni e di attivare i suoi meccanismi di autodifesa.
Ma perché i meccanismi di autodifesa rispondono in modo diverso alla malattia a seconda della varietà? Questa è una delle domande che ha spinto il team del Progetto Ring (ACT190001) a identificare i geni coinvolti nella risposta delle varietà Santina e Bing, che presentano una diversa suscettibilità alla malattia, essendo Santina tollerante e Bing suscettibile.
Grazie alle ricerche condotte, sono stati compiuti progressi nella comprensione di questa risposta differenziata alla Pss, sulla base dei geni differenzialmente espressi (DEG) nel tempo, cioè dal momento dell'inoculazione della pianta con il batterio fino alla manifestazione dei primi sintomi, svelando le vie e i processi coinvolti.
Ciò potrebbe essere fatto studiando il profilo trascrittomico, cioè i DEG, ottenendo le sequenze degli RNA sintetizzati dalle piante in risposta agli stimoli applicati. La trascrittomica è un potente strumento offerto dalla biologia molecolare in quanto permette, in qualsiasi momento, di conoscere l'espressione dei geni nei tessuti vegetali.
In altre parole, con lo studio del trascrittoma è possibile scoprire quale meccanismo o quali meccanismi vengono attivati nella pianta affinché possa tollerare una situazione di stress. D'altra parte, queste analisi permettono di stimare l'intensità di questi segnali, il che consente di quantificare il livello della risposta di difesa e di metterlo in relazione con i cambiamenti anatomici osservati.
In termini generali, si sa che il sistema di difesa delle piante coinvolge diversi tipi di meccanismi che interagiscono tra loro per far fronte all'attacco di un determinato patogeno, come le caratteristiche specifiche della varietà, la "pattern-activated immunity" (PTI) e la "effector-activated immunity" (ETI) (Figura 1).
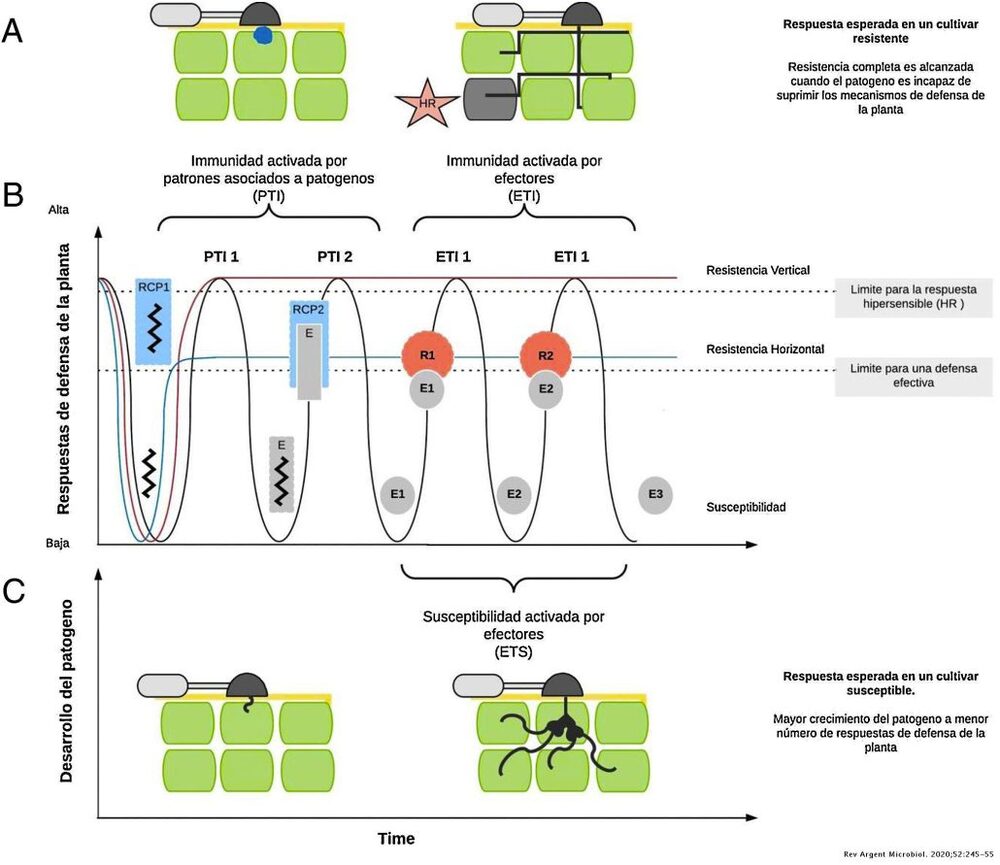 Figura 1: Schema dell'immunità attivata dal patogeno (PTI) e dell'immunità attivata dagli effettori (ETI). A) Risposta completa al patogeno che ne impedisce l'ingresso o l'avanzamento attraverso la risposta di ipersensibilità (HR), massima espressione di resistenza al patogeno. B) Diagramma delle risposte di difesa della pianta nel tempo. La curva rossa descrive la risposta attesa in un processo di resistenza verticale, che è una resistenza specifica codificata da un solo gene, mentre la curva blu descrive la risposta attesa durante la resistenza orizzontale, quella codificata da più geni, o poligenica. C) Infezione riuscita di un patogeno in un ospite suscettibile (Burbano-Figueroa, 2020).
Figura 1: Schema dell'immunità attivata dal patogeno (PTI) e dell'immunità attivata dagli effettori (ETI). A) Risposta completa al patogeno che ne impedisce l'ingresso o l'avanzamento attraverso la risposta di ipersensibilità (HR), massima espressione di resistenza al patogeno. B) Diagramma delle risposte di difesa della pianta nel tempo. La curva rossa descrive la risposta attesa in un processo di resistenza verticale, che è una resistenza specifica codificata da un solo gene, mentre la curva blu descrive la risposta attesa durante la resistenza orizzontale, quella codificata da più geni, o poligenica. C) Infezione riuscita di un patogeno in un ospite suscettibile (Burbano-Figueroa, 2020).
Tuttavia, non è ancora noto come questi meccanismi funzionino in modo specifico per ogni varietà di ciliegio. Alcuni dei fattori che influenzano la suscettibilità alla Pss sono: il momento della caduta delle foglie, la rottura della dormienza e la generazione di fellogeno attivo, la data di fioritura, l'età della pianta, la genetica della pianta e il portainnesto utilizzato.
C'è anche poca chiarezza sul modo in cui il ciliegio rileva la presenza di patogeni per mezzo di "pattern molecolari specifici associati ai patogeni" (PAMP), o sul modo in cui evita i danni da patogeni e si difende rilasciando i cosiddetti "pattern molecolari associati ai danni" (DAMP).
Infine, le piante possono rilevare gli effettori secreti dal patogeno, proteine che interferiscono con il metabolismo della pianta allo scopo di bloccare la sua risposta di difesa PTI, attivando così l'ETI.
Alla ricerca di queste risposte, è stato effettuato un saggio di infezione in condizioni controllate utilizzando piante di 1 anno delle varietà Bing e Santina, innestate su Gisela 12, considerando come controllo acqua distillata sterile. È stato effettuato un taglio obliquo a 10-15 cm dall'apice e sono stati inoculati 20 µL di una sospensione batterica di 108 CFU/mL con il ceppo Pss11116B1, altamente virulento.
Le piante sono state mantenute in condizioni di serra con temperature tra i 25 e i 27°C e umidità tra il 57-67%; sono stati prelevati 2 campioni di tessuto ed è stato estratto l'RNA per l'analisi del trascrittoma, il primo a 1 giorno post-inoculo (dpi) per analizzare la risposta precoce e il secondo a 7 dpi per analizzare come si attiva la difesa in presenza di sintomi.
Successivamente, questi campioni sono stati inviati per il sequenziamento, effettuato con la piattaforma Illumina (RNAseq) e da questi dati è stato ottenuto il profilo trascrittomico per analizzare la differenza tra i diversi profili DEG.
I risultati
A 5 dpi sono stati osservati sintomi di secrezione di gomma e necrosi intorno alla zona di inoculo in entrambe le varietà, mentre le piante di controllo non hanno mostrato cambiamenti, tranne la cicatrizzazione della ferita (Figura 2).
 Figura 2: Tipi di sintomi nelle giovani piante infettate con il ceppo Pss11116B1 e il controllo a 7 dpi. A) Guarigione della ferita nella pianta di controllo con acqua sterile. B-C-D) Necrosi intorno alla zona di inoculo e inizio della produzione di gommosi.
Figura 2: Tipi di sintomi nelle giovani piante infettate con il ceppo Pss11116B1 e il controllo a 7 dpi. A) Guarigione della ferita nella pianta di controllo con acqua sterile. B-C-D) Necrosi intorno alla zona di inoculo e inizio della produzione di gommosi.
I risultati dell'analisi trascrittomica a 1 dpi hanno rivelato che la varietà Santina, meno suscettibile all'infezione da Pss, attiva la sua risposta più rapidamente, esprimendo in modo differenziato il 70% di geni in più rispetto a Bing (831 DEG). È stato inoltre osservato che le piante della varietà Bing hanno rafforzato gradualmente la loro risposta, raggiungendo, a 7 dpi, una risposta più robusta (1471 DEG). L'analisi dettagliata di questi DEG fornirà informazioni sulle risposte comuni e specifiche della varietà.
Lo studio dettagliato del profilo trascrittomico di questi ceppi evidenzia il numero di processi che, in termini semplici, sono attivi nei campioni di Santina. La Figura 3 elenca questi processi dopo 1 dpi; la dimensione della bolla (cerchio) indica il numero di geni differenzialmente espressi in ciascun processo. È evidente come in Santina il profilo trascrittomico includa un numero maggiore di processi (sia attivati che repressi) rispetto a Bing.
 Figura 3: Analisi dell'arricchimento in risposta all'inoculazione con Pss 11116B1. A) Processi in Santina. B) Processi in Bing. La dimensione della bolla indica il numero di geni associati a ciascun processo. L'asse delle X rappresenta l'arricchimento, un numero più alto indica una maggiore rilevanza del processo. Il colore indica la significatività; quando il colore diventa rosso, esprime una maggiore significatività statistica.
Figura 3: Analisi dell'arricchimento in risposta all'inoculazione con Pss 11116B1. A) Processi in Santina. B) Processi in Bing. La dimensione della bolla indica il numero di geni associati a ciascun processo. L'asse delle X rappresenta l'arricchimento, un numero più alto indica una maggiore rilevanza del processo. Il colore indica la significatività; quando il colore diventa rosso, esprime una maggiore significatività statistica.
Tra questi, spicca la differenza nella sovraespressione dei geni di rimodellamento della parete cellulare. La parete cellulare è una rete complessa e dinamica che, tra le altre funzioni, fornisce protezione alle piante agendo come una barriera fisica che impedisce o ostacola l'ingresso di agenti patogeni. Nel Bing questa sovraespressione si osserva a 7 dpi.
Un altro dato rilevante è che i geni legati al processo fotosintetico sono meno espressi nella varietà Santina, il che potrebbe essere interpretato come un reindirizzamento delle risorse della pianta per promuovere le risposte di difesa.
Questa risposta è completata dall'analisi dei geni di risposta alla difesa differenzialmente espressi; nella figura si può notare che Santina esprime precocemente un maggior numero di geni di difesa rispetto a Bing, tra cui quelli coinvolti nella risposta di difesa contro i batteri sono sovraespressi, mentre in Bing questi sono attivi solo a 7 dpi.
In conclusione, il momento della risposta è cruciale, la varietà Bing impiega più tempo per attivare i suoi meccanismi di difesa contro la Pss (Figura 4), il che può spiegare la sua maggiore suscettibilità alla malattia; mentre Santina è in grado di rispondere precocemente e con un maggior numero di processi coinvolti nella modulazione della difesa.
 Figura 4: Numero di geni di risposta alla difesa in Santina e Bing inoculate con Pss 11116B1 nel tempo.
Figura 4: Numero di geni di risposta alla difesa in Santina e Bing inoculate con Pss 11116B1 nel tempo.
Sebbene la strada da percorrere per comprendere appieno i meccanismi di interazione pianta-patogeno sia ancora lunga, questo studio ha permesso di compiere i primi passi che consentono di visualizzare come il controllo della Pss nel ciliegio potrebbe basarsi sul potenziamento dei processi di autodifesa della pianta stessa.
Grazie a queste scoperte, è possibile che molto presto saranno disponibili induttori di resistenza efficaci e a basso costo per ridurre i danni causati dalla Pss nel ciliegio, riducendo l'uso del rame e limitando la comparsa di ceppi batterici ad esso resistenti, evitando così danni alla salute umana e all'inquinamento ambientale.
Fonte: Redagrícola
Immagini: Redagrícola
Cherry Times - Tutti i diritti riservati








