Bacterial cancer is one of the main diseases affecting cherry, which is associated with a complex of bacteria of the genus Pseudomonas, being Pseudomonas syringae pv. syringae (Pss) the main causal agent of the disease in Chile. It causes damage at all stages of the plant, causing death of young plants in the nursery and yield reduction in adult plants that can reach 50% of the production, and a marked reduction in the useful life of the orchard.
The factors involved in the development of the disease are multiple and intertwine to trigger the infection and favour the subsequent establishment of the bacterial complex in different organs of the plant. These include: age of the plant, degree of virulence of the bacterial strains, environmental conditions, presence of other infectious agents, action of abiotic stresses such as water and, finally, the variety used.
If to these we add that today the curative control of bacterial cancer is mainly based on repeated treatments with copper products and that the bacterium is capable of acquiring and expressing resistance mechanisms and generating strains that are increasingly tolerant to copper, then it is a priority to focus control through strategies to prevent infection and reduce the spread of the bacterium.
Currently, to limit the spread of the bacterium in the orchard, it is advised to remove affected branches and cankers. Spread can occur through the use of infected plant material (e.g. by grafting), rain, wind and contaminated handling tools (e.g. during pruning).
Pss enters the plant through natural openings such as stomata and lenticels, or artificial openings such as wounds caused by leaf fall, growth cracks, wounds caused by the conduction system or micro-infections generated by the bacterium's ability to freeze water (ice nucleation) at temperatures just above 0°C.
Differential varietal susceptibility
Regarding the export of cherries, the main varieties produced and exported in Chile during the 2022/23 season were, in order of importance: Lapins, Santina, Regina and Bing (Asoex, 2023). However, according to observations of symptoms in commercial orchards and laboratory infection tests, Bing is the variety with the highest susceptibility to bacterial cancer, followed by Regina, Lapins and, lastly, Santina.
It is important to consider that the degree of differential susceptibility to bacterial canker present in the different cherry varieties is primarily related to the plant's ability to rapidly detect the presence of the pathogen in internal tissues and activate its self-defence mechanisms.
But why do self-defence mechanisms respond differently to disease depending on the variety? This is one of the questions that motivated the Ring Project team (ACT190001) to identify the genes involved in the response of the Santina and Bing varieties, with contrasting susceptibility to the disease, Santina being tolerant and Bing susceptible.
Through the research carried out, progress was made in understanding this differential response to Pss, based on the differentially expressed genes (DEG) over time, that is, from the time of inoculation of the plant with the bacterium to the manifestation of the first symptoms, unravelling the routes and processes involved.
This could be done by studying the transcriptomic profile, i.e. the DEGs, obtaining the sequences of the RNAs synthesised by the plants in response to the stimuli applied. Transcriptomics is a powerful tool offered by molecular biology as it allows, at any given moment, to know the expression of genes in plant tissues.
In other words, with a transcriptome study it is possible to find out which mechanism or mechanisms are being activated in the plant in order for it to tolerate a stress situation.On the other hand, these analyses make it possible to estimate the intensity of these signals, which makes it possible to quantify the level of the defence response and relate it to the anatomical changes observed.
In general terms, what is known about the plant defence system is that it involves different types of mechanisms that interact with each other to cope with the attack of a given pathogen, such as variety-specific characteristics, "pattern-activated immunity" (PTI) and "effector-activated immunity" (ETI) (Figure 1).
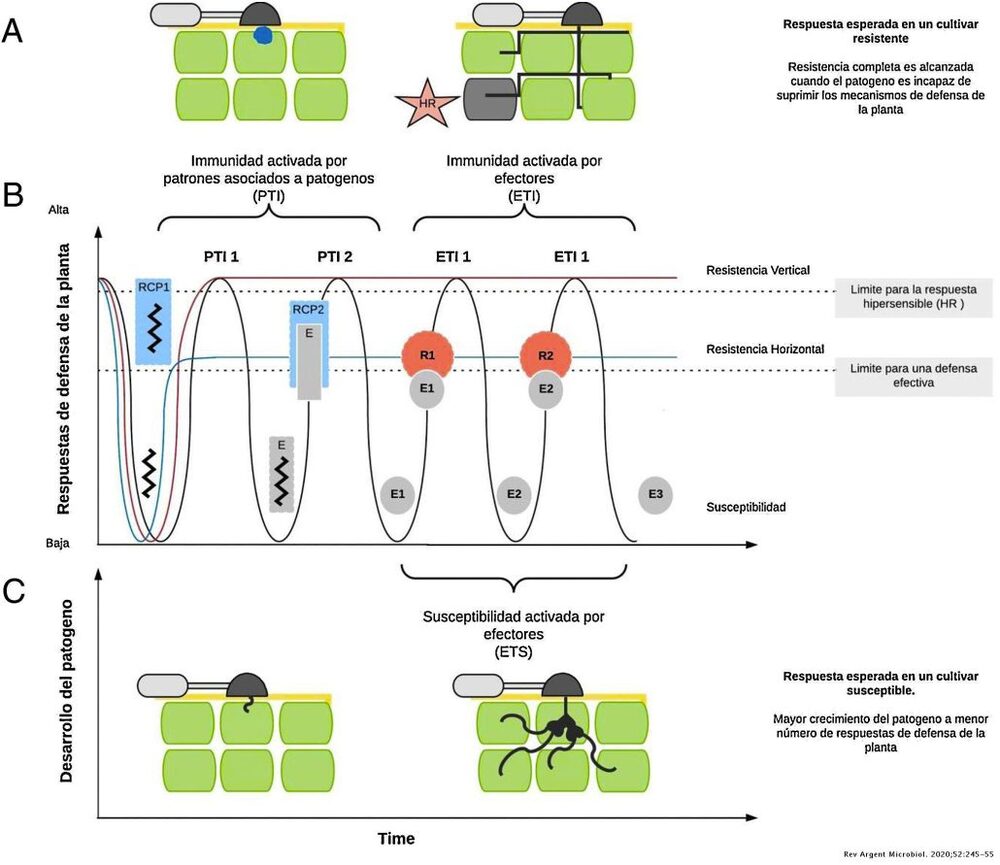
Image 1: Scheme of pathogen-associated pattern-activated immunity (PTI) and effector-activated immunity (ETI). A. Complete response to the pathogen preventing its entry or advance through the hypersensitive response (HR), the maximum expression of resistance to the pathogen. B) Diagram of plant defence responses over time. The red curve describes the expected response in a vertical resistance process, which is a specific resistance encoded by one gene, and the blue curve describes the expected response during horizontal resistance, that encoded by several genes, or polygenic. C) Successful infection of a pathogen in a susceptible host (Burbano-Figueroa, 2020)
However, it is not yet known how these mechanisms function in a variety-specific manner in each cherry variety. Certainly, some of the factors, which affect susceptibility to Pss, are: timing of leaf fall, dormancy break and generation of active phellogen, flowering date, plant age, plant genetics and rootstock used.
There is also little clarity on how the cherry tree detects the presence of pathogens by means of "pathogen-associated specific molecular patterns" (PAMPs), or how it avoids pathogen damage and defends itself by releasing what are known as "damage-associated molecular patterns" (DAMPs).
Finally, plants can detect effectors secreted by the pathogen, which are proteins that interfere with the plant's metabolism with the aim of blocking its PTI defence response, thus activating the ETI.
In search of these responses, an infection test was carried out under controlled conditions using 1-year-old plants of Bing and Santina varieties, grafted on Gisela 12, considering sterile distilled water as a control. A bevel cut was made 10-15 cm from the apex and 20 µL of a bacterial suspension of 108 CFU/mL was inoculated with the highly virulent Pss strain Pss11116B1.
Plants were maintained in greenhouse conditions with temperatures between 25 and 27°C and humidity between 57-67%, 2 tissue samples were taken and RNA was extracted for transcriptome analysis, the first one at 1 day post inoculation (dpi) to analyse the early response and the second one at 7 dpi to analyse how the defence is activated in the presence of symptoms.
Subsequently, these samples were sent for sequencing, which was done using the Illumina platform (RNAseq) and from these data the transcriptomic profile was obtained to analyse the difference between the different DEG profiles.
Results
At 5 dpi, symptoms of gum secretion and necrosis around the inoculation zone were observed in both varieties, while the control plants showed no changes, except for wound healing (Figure 2).

Image 2: Types of symptoms in young plants infected with strain Pss11116B1 and control at 7 dpi. A) Wound healing in control plant with sterile water. B-C-D) Necrosis around the inoculation zone and start of gum production.
The results of the transcriptomic analysis at 1 dpi revealed that the Santina variety, which is less susceptible to Pss infection, activates its response more quickly, differentially expressing 70% more genes than Bing (831 DEG). It was also observed that the plants of the Bing variety gradually strengthened their response, reaching, at 7 dpi, a more robust response (1471 DEG). Detailed analysis of these DEGs will provide insight into common and variety-specific responses.
Studying the transcriptomic profile of these strains in detail highlights the number of processes that, in simple terms, are active in the Santina samples. Figure 3 lists these processes after 1 dpi, the size of the bubble (circle) indicates the number of differentially expressed genes in each process. It is evident how in Santina the transcriptomic profile includes a higher number of processes (both activated and repressed) compared to Bing.

Image 3: Enrichment analysis in response to inoculation with Pss 11116B1. A) Processes in Santina. B) Processes in Bing. The size of the bubble indicates the number of genes associated with each process. The X-axis represents the enrichment, a higher number indicates higher relevance of the process. The colour indicates significance, as the colour turns red it expresses a higher statistical significance.
Among them, the difference in the overexpression of cell wall remodelling genes stands out. The cell wall is a complex and dynamic network that, among other functions, provides protection to plants by acting as a physical barrier that prevents or hinders the entry of pathogens. In Bing this overexpression is observed at 7 dpi.
Another relevant finding is that genes linked to the photosynthetic process are less expressed in the Santina variety, which could be interpreted as a redirection of plant resources to promote defence responses.
This response is complemented by analysing the differentially expressed defence response genes, in the figure it can be seen that Santina expresses a greater number of defence genes early than Bing, among which those involved in the defence response against bacteria are overexpressed, while in Bing these are only active at 7 dpi.
In conclusion, the time of response is crucial, the Bing variety takes longer to activate its defence machinery against Pss (Figure 4) which may explain its higher susceptibility to disease; while Santina is able to respond early and with a higher number of processes involved in modulating its defence.

Image 4: Number of defence response genes in Santina and Bing inoculated with Pss 11116B1 over time.
Although there is still a long way to go to fully understand the mechanisms of plant-pathogen interaction, this study has made it possible to take the first steps that allow us to visualise how the control of Pss in cherry could be based on enhancing the plant's own self-defence processes.
Thanks to these discoveries, it is possible that very soon effective and low-cost resistance inducers will be available to reduce the damage caused by Pss in cherry trees, reducing the use of copper and limiting the appearance of bacterial strains resistant to it, thus avoiding damage to human health and environmental pollution.
Source: Redagrícola
Images: Redagrícola
Cherry Times - All rights reserved








