The post-harvest phase is a critical period that determines the productive success of a cherry orchard for the following season, where the accumulation of carbon and nitrogen reserves plays a key role.
This accumulation can be reduced due to various climatic factors typical of the summer season in central Chile, such as high maximum temperatures, intense solar radiation, high water demand, and low relative humidity. These factors can easily create stress conditions in plants. During this period, from December to March, it is crucial to have stress-free plants and ensure efficient reserve accumulation, which will serve as the engine to support flowering and bud break in the following spring.
1. Understanding the reserve content in cherry trees
The cherry tree is a species with a short production period, and harvesting can start 50 to 75 days after full bloom, depending on the variety.
The flowering will depend, in turn, on the nutritional reserves stored in tissues such as roots, buds, and wood. Additionally, since root growth activation in spring can begin between 45 and 60 days after full bloom when soil temperature reaches at least 15°C, the accumulation of reserves will be critical. This applies both to the fruit setting processes and the initial fruit growth phase (active cell division), which can directly impact final fruit size and, therefore, yield.
 Image 1.
Image 1.
It is therefore important to quantify or have a reference for the accumulated reserves during the post-harvest period. Currently, the most commonly used analyses in the industry to determine reserve accumulation are the arginine content in roots and starch in buds or floral structures.
Regarding the impact of reserve accumulation on production parameters, trials conducted in previous seasons by CER have demonstrated a positive relationship between reserve content, measured as arginine content in roots, and yield. This relationship was more evident in Lapins than in Bing, showing a varietal response associated with the variety's productive potential.
 Image 2: Linear regressions for variables arginine concentration in roots and yield in Lapins and Bing cherry trees.
Image 2: Linear regressions for variables arginine concentration in roots and yield in Lapins and Bing cherry trees.
2. Factors influencing reserve accumulation
 Image 3.
Image 3.
Reserves begin to accumulate significantly from harvest until leaf fall, when the highest concentration occurs in the woody organs of the cherry tree. The accumulation of carbon and nitrogen reserves mainly depends on temperature, nutritional conditions, and the plant's water status.
For carbon reserves, these directly depend on the plant's photosynthetic efficiency, which in turn is influenced by climatic conditions and the various agronomic practices carried out during this period.
In the case of nitrogen reserves, it is crucial to have adequate irrigation and fertilization, which keep plants in optimal condition during the months of higher evapotranspirative demand and counteract the effects of high temperatures and solar radiation, which have increased in recent years due to climate change. In fact, most production reductions do not occur at extreme temperatures but at relatively moderate temperatures (between 34°C and 38°C), which can reduce net photosynthesis to nearly zero.
International studies have shown that temperatures above 38°C deactivate the enzymes involved in photosynthesis. In this scenario, it is essential to integrate traditional management with strategies to reduce heat stress in plants, thereby maximizing photosynthetic and productive potential.
3. Management to mitigate heat stress in cherry trees
3.1. Sunblocks
In the last decade, sunblocks have been widely used to mitigate the effects of temperature and summer radiation on fruit tree production, both pre-harvest and post-harvest. There is a wide range of these products, which can be composed of kaolinite, silicates, or fatty acids such as phospholipids or oligosaccharides (colorless products). They are used during months of high temperatures and solar radiation (from December to February), as they can mitigate heat stress by lowering leaf temperatures.
This protection enables greater absorption of carbon from the atmosphere into the plant by reducing stomatal closure. The effect of sunblocks has been extensively studied in apple production, which can experience severe sun damage on fruits and leaves depending on location and variety. In cherry trees, sunblocks are frequently used, especially colorless ones pre-harvest, while both colorless and kaolinite-based products are used post-harvest.
 Image 4: High temperature damage in Santina cherry trees in central Chile, January 2023.
Image 4: High temperature damage in Santina cherry trees in central Chile, January 2023.
In terms of mode of action, the application of sunblocks reflects excess radiation, including photosynthetically active radiation (PAR), ultraviolet (UV), and infrared (IR), reducing the risk of sunburn damage to leaves and fruits, preserving tissue integrity, and preventing chronic photoinhibition after a heat stress event.
Studies conducted by CER on post-harvest cherry trees highlighted a higher presence of senescent leaves in control plants without application compared to plants treated with sunblocks (colorless or kaolinite-based), demonstrating the direct effect of such products on the most exposed leaf mass.
 Image 5: Plant status on March 3, 2022, 21 days after the second application of sunblocks on Regina cherry trees, 2021-2022 season.
Image 5: Plant status on March 3, 2022, 21 days after the second application of sunblocks on Regina cherry trees, 2021-2022 season.
Other studies show that pomegranate plants treated with a sunscreen based on kaolin achieve a reduction in leaf temperature by 2.5°C on average, while the control treatment records a leaf temperature equivalent to 38.1°C. This data is particularly significant considering that the reduction in leaf temperature is achieved at low levels, where Rubisco undergoes deactivation, as indicated in the previous paragraphs.
Similar results have been observed in trials on cherry trees treated with sunscreens, where a decrease in leaf temperature of 1-2°C was observed compared to an untreated control, with a significant effect on stomatal opening.
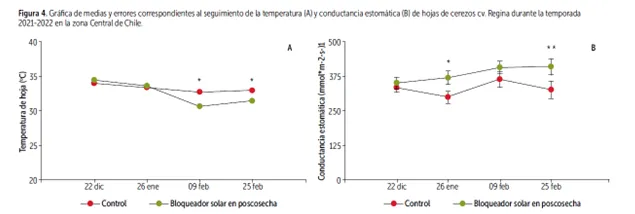 Image 6: Graph of averages and corresponding errors for monitoring temperature (A) and stomatal conductance (B) of Regina cherry tree leaves during the 2021-2022 season in central Chile.
Image 6: Graph of averages and corresponding errors for monitoring temperature (A) and stomatal conductance (B) of Regina cherry tree leaves during the 2021-2022 season in central Chile.
Furthermore, significant differences were observed in the stomatal conductance of Regina plants treated with kaolinite-based and colorless sunscreens, compared to untreated plants. Regardless of the type of sunscreen applied, the treated plants did not fall below 200 mmol*m-2s-1, indicating a moderate level of stress, unlike the untreated control plants.
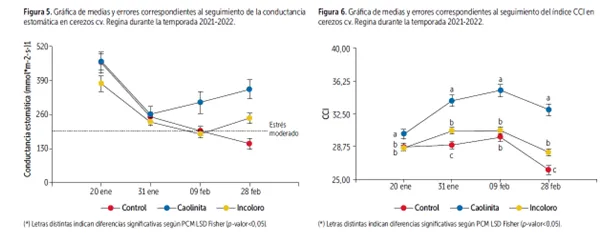 Image 7: Graph 1: Graph of averages and corresponding errors for monitoring stomatal conductance in Regina cherries during the 2021-2022 season. Graph 2: Graph of averages and corresponding errors for monitoring the CCI index in Regina cherries during the 2021-2022 season.
Image 7: Graph 1: Graph of averages and corresponding errors for monitoring stomatal conductance in Regina cherries during the 2021-2022 season. Graph 2: Graph of averages and corresponding errors for monitoring the CCI index in Regina cherries during the 2021-2022 season.
Another variable to consider in determining the physiological status of plants under biotic and/or abiotic stress is the chlorophyll content, responsible for the green coloration of leaves. This is because plants can modify their internal concentration, reflecting changes in plant metabolism in response to these events.
The chlorophyll content index (CCI) in leaves, measured with a portable device (MC-100, Apogee Instruments), was found to be significantly higher in plants treated post-harvest with kaolinite-based sunscreens, statistically differing from plants treated with colorless sunscreens and untreated plants. These treated plants remained greener, more functional, and accumulated reserves for a longer period. The age of the leaves and their physiological state are key factors determining chlorophyll content.
3.2. Application of Biostimulants
The complementary use of nutritional correctors and/or biostimulants during critical periods of cultivation has recently gained importance as a strategy to mitigate stress in plants. Biostimulants, according to the most accepted definition, are substances whose primary function is to stimulate the biochemical processes of the plant to enhance nutrient absorption and efficiency, tolerance to abiotic stresses, and yield improvement. This category includes products with various formulations based on humic and fulvic acids, proteins, biopolymers, silicates, seaweed extracts, among others.
 Image 8.
Image 8.
Regarding stress mitigation, seaweed-based biostimulants have been the most commonly used in the post-harvest period. These extracts naturally contain polysaccharides, macro- and micronutrients, amino acids, and phytohormones, among others, which vary depending on the type of seaweed and the extraction method used to obtain them.
The most commonly used seaweeds for this purpose are extracts based on Ascophyllum nodosum and Ecklonia maxima, whose nutritional composition triggers a series of physiological responses in the plant, increasing its tolerance to abiotic stresses, whether thermal, water, or saline.
Although their mechanism of action is not fully understood, some studies have shown that their application induces changes in the metabolic pathways of the plant's own hormones, altering the endogenous synthesis of auxins, cytokinins, and abscisic acid, in addition to generating synergistic effects between their components. This allows plants to optimize their physiological processes, making them better prepared nutritionally to face stress events and maintain their productive potential.
Studies conducted by CER have shown that the application of seaweed-based biostimulants in post-harvest programs for cherry trees impacts nitrogen reserve accumulation in the roots, evaluated during the winter dormancy period.
Regarding E. maxima-based biostimulants, late applications (March-April) have been shown to increase carbon reserves in the buds, providing a significant contribution compared to untreated controls. Similarly, early post-harvest applications (January-February) of seaweed extracts from the genus A. nodosum significantly increase reserve accumulation, measured as arginine concentration in the roots.
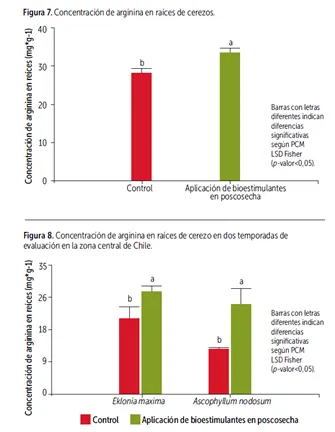 Image 9: Graph 1: Arginine concentration in cherry tree roots. Graph 2: Arginine concentration in cherry tree roots during two evaluation seasons in central Chile.
Image 9: Graph 1: Arginine concentration in cherry tree roots. Graph 2: Arginine concentration in cherry tree roots during two evaluation seasons in central Chile.
4. Final Considerations
 Image 10.
Image 10.
In conclusion, proper irrigation and fertilization management in post-harvest, integrated with sunscreen and/or algae-based biostimulant application strategies to mitigate thermal stress caused by high summer temperatures, elicits a plant response that results in greater reserve accumulation, thus ensuring a strong start to the next season.
The general recommendation is to apply proper management that results in greater reserve accumulation, ensuring a good start to the next season. In this sense, the general recommendation is to apply sunscreens, either kaolinite-based or colorless, as early as possible after harvest and, complementarily, seaweed-based biostimulants every 15-20 days throughout the period of high temperatures.
The frequency of these applications will depend on the characteristics of the orchard (climate, latitude, vigor, phytosanitary status of the plants, etc.) and may be more frequent in areas with higher temperatures or where plants are more susceptible to thermal stress.
Source: Redagrícola
Images: Millanao et al., 2024; SL Fruit Service
Marjorie Millanao B., Fernando Quezada M., Carolina Concha E.
DEPARTAMENTO PRODUCCIÓN AGRÍCOLA CER - Chile
Cherry Times - All rights reserved








