“Health is the right balance between hot and cold.” A good budbreak of broadleaf trees is the result of exposure to a certain amount of winter cold and spring warmth. However, in recent seasons, this combination has not occurred to the required extent, negatively impacting the phenology of fruit trees.
As a result of anticipated climate change, various production areas around the world, as well as in central-southern Chile, have experienced colder than usual winters. This has sparked interest in applying models based on the concept of cold/heat interaction to explain the effects of these changes on the behavior of fruit trees.
Extreme climate variations in recent years, due to natural cycles such as the El Niño Southern Oscillation and the effects of climate change, have generated uncertainty about the behavior of trees in every season.
More than ten years ago, it was predicted that one of the effects of climate change on fruit growing would be a reduced availability of cold in winter in various production areas, with a negative impact on production to the extent that exposure to cold would not be sufficient to meet the minimum requirements of a specific fruit species, i.e., its chilling requirement.
The Three Phases of Dormancy
During winter, deciduous trees lose their leaves and enter a state of dormancy. In this state, they cease growth as a survival mechanism against unfavorable winter conditions. This is a continuous process in which three phases occur sequentially.
In the first phase, trees prepare for dormancy: they respond to shorter days and low temperatures, mobilizing nutrients and other compounds, including inhibitors like abscisic acid (ABA), from the leaves to the twigs.
The cessation of photosynthetic activity, the polymerization of sugars, the blockage of transport, and the accumulation of inhibitors like ABA, leading to leaf drop, mark the beginning of the endodormancy phase or deep dormancy. In this phase, there is no growth due to the internal conditions of the buds, which therefore will not sprout even in favorable environments.
Exposure to winter conditions degrades the inhibitors, their synthesis decreases, and the bud enters eco-dormancy, meaning it has the capacity to sprout but will not do so as long as winter conditions are maintained. Buds will grow when days are warmer and brighter in spring.
The way to monitor the recession trend is to quantify winter cold. For this purpose, several models are available, the most commonly used being: Chilling Hours below 7°C, Richardson or Utah Chilling Units, and Portions of chilling according to the dynamic model.
Species and cultivars have different cold thresholds to achieve sprouting capability. Therefore, they need a heat accumulation, which can be quantified with models such as Growing Degrees (GD) and Decline Degree Hours (GDH).
Cold-Heat Model
Before the turn of the century, the amounts of cold and heat post-harvest for deciduous fruit trees were expressed as successive and independent events. In the 2000s, researcher Eduardo Tersoglio from INTA Mendoza demonstrated that the buds of Bing cherries with greater exposure to winter cold required less heat to sprout.
In 2010, researchers from the U.S. Forest Service, led by Connie Harrington, proposed a model of interaction or trade-off between the amount of cold and heat exposure for the emergence from dormancy of a conifer species. This model was later applied to fruit trees by the research group led by Professor Ted DeJong in California and published in 2014 (Figure 1).
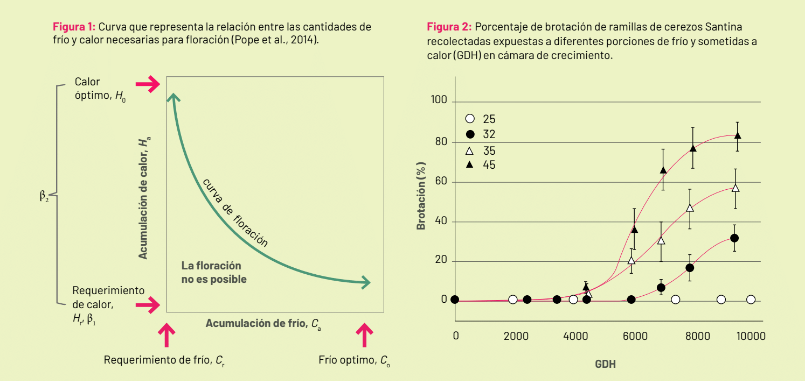 Image 1: Curve representing the relationship between the amounts of cold and heat necessary for flowering (Pope et al., 2014). Image 2: Percentage of germination of Santina cherry branches collected exposed to different cold portions and subjected to heat (GDH) in the growth chamber.
Image 1: Curve representing the relationship between the amounts of cold and heat necessary for flowering (Pope et al., 2014). Image 2: Percentage of germination of Santina cherry branches collected exposed to different cold portions and subjected to heat (GDH) in the growth chamber.
The model indicates that, with increased exposure to cold during endodormancy, buds increase their capacity to sprout, which is reached with subsequent exposure to heat. However, the more buds are exposed to cold, the less heat they require to sprout.
Assuming that the transition between endo and eco-dormancy occurs when 50% of the buds are capable of sprouting, two key parameters of the process can be recognized in this interaction: the cold and the heat required.
The minimum amount of cold for 50% of the buds to sprout without thermal limitations (with maximum heat) is the chilling requirement. On the other hand, the minimum amount of heat to sprout 50% with the maximum amount of cold would be the heat requirement.
The cold/heat interaction model allows for understanding dormancy as a continuous and dynamic process. Furthermore, it suggests a compensation in phenological progress. In other words, a warm spring will compensate for a potentially low accumulation of cold in winter and vice versa.
With the support of FIC Maule and using the technique of forced sprouting in growth chambers, the chilling and heat requirements of newly introduced cherry and apple cultivars are being determined (Photo 2). For quantification, chilling portions and GDH have been chosen, in line with the preference of most experts for these models.
It is then possible to construct response curves to heat for branches collected with different amounts of winter cold, with varying exposure to cold (Figure 2). With the amounts of cold and heat for 50% sprouting, it is possible to fit a curve with which to estimate the heat and chilling requirements for each cultivar. Although the most determining factor for the value of the chilling requirement is given by the species and cultivar, it also varies based on the tree’s vigor (rootstock, age).
In the orchard, it is necessary to establish the timing of entry into endodormancy to begin counting the cold and compare the actual cold accumulation with the chilling requirement of each cultivar. In apple trees, it has been established and measured that the onset of endodormancy occurs when the tree shows a drop of 50% of its leaves.
In cherry trees, there are various positions to set the start of cold counting. It is perhaps less relevant than in apples, where both leafless cultivars and unharvested cultivars in autumn can be found. The entry into dormancy of cherry trees, manifested by leaf drop, is regulated by the shortening of days and low temperatures.
Using data from two seasons and four orchards in the Maule region, the heat and cold curves of Santina and Lapins have been fitted (Figure 3). Although a more approximate fit can be obtained with a larger number of data points, the trends plotted allow for estimating the behavior of both cultivars.
Santina, despite showing similar chilling requirements to Lapins in endodormancy, has a greater need for heat to sprout when cold is abundant. Therefore, Santina will express its potential for phenological progress in areas with warm springs. Lapins can be grown in places with lower heat accumulation in spring.
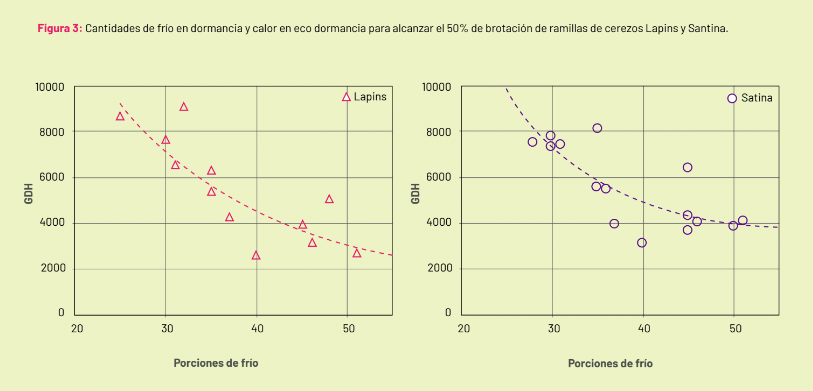 Image 2: Response curve to heat of Santina and Lapins cherry branches exposed to different amounts of cold, after 2016-2017 and 2017-2018 seasons. Each color indicates the average of branches with the specified amount of cold. To estimate the amount of heat necessary for sprouting, branches collected in mid-spring were subjected to different heat exposures.
Image 2: Response curve to heat of Santina and Lapins cherry branches exposed to different amounts of cold, after 2016-2017 and 2017-2018 seasons. Each color indicates the average of branches with the specified amount of cold. To estimate the amount of heat necessary for sprouting, branches collected in mid-spring were subjected to different heat exposures.
This research allows for advancing and adapting cultural practices based on understanding the behavior of each cultivar, such as adjusting pruning dates to favor new growth. For example, budbreak in warm areas, when sprouting is expected to occur quickly, may be favored by delaying pruning for the same reason. On the contrary, in places with low heat, pruning can be advanced to stimulate growth.
We hope to present the results of our ongoing research to improve practices in the cherry industry and optimize performance in the next few seasons.
Use of Canopies
In recent seasons, weather conditions have been contrasting: the winter of 2023 had limited cold accumulation, and spring remained cold until December. The worst combination occurred, as there was no compensation between cold and heat to normalize phenological progress.
Cherry trees in sheds with some environmental modification systems, such as shade nets, tunnels, or canopies with perimeter closures, which favored thermal accumulation in spring, recorded better production data.
In 2024, with more typical winter conditions (cloudy days and rain until June, low temperatures, and fog), environmental modification systems did not make a difference and sometimes, on the contrary, were counterproductive. At low temperatures, trees under shade nets can be slightly warmer than the outside.
Similarly, plastic canopies used in spring do not make a difference, as they have a high cold accumulation in winter, and without this input, their use can be questioned due to the reduction of solar radiation they generate.
Source: Mundoagro
Images: Mundoagro
Álvaro Sepúlveda, Arene di Loreto, Daniela Simeone, Maurizio Fuentes
Cherry Times - All rights reserved








