The Chilean cherry is experiencing its most complex moment: after a decade of expansion, the last season was marked by a decline in selling prices. Today, the industry’s competitiveness requires fruit of good size, firm, in excellent condition, and with maximum use of available resources.
In this scenario, calcium remains a strategic ally in the fruit’s physiology, as it is crucial for firmness and postharvest life.
Over the last decade, the cherry tree has become the most relevant fruit crop in Chilean horticulture, reaching more than 76,000 hectares planted (ODEPA, 2025) and with a marked dependence on exports to China, which accounts for over 90% of shipments (Muñoz, 2025).
However, in the last season, grower returns suffered a sharp decline due to relative oversupply and the growing demand from the market for high-quality fruit.
Improving efficiency and quality
This is a scenario where it becomes essential to improve efficiency in the use of resources while ensuring quality attributes that allow product differentiation and the maintenance of competitive prices.
Among the main attributes, flesh firmness is crucial, both for consumer preference and for its close relationship with resistance to mechanical damage, tolerance to long-distance transport, and the incidence of postharvest rots.
Fruit firmness is largely determined by the structure and dynamics of the cell wall, where calcium plays a fundamental role.
This cation not only participates in essential physiological processes such as signaling and membrane integrity but also stabilizes pectins in the middle lamella through the formation of “egg-box” bonds, which provide rigidity and reduce the loss of cellular cohesion.
Benefits of soil applications
Consequently, an adequate accumulation of calcium in the fruit can delay softening, extend shelf life, and increase the share of exportable fruit.
Although numerous studies have been conducted on foliar applications or postharvest dips with calcium-rich solutions, the results have proven inconsistent and, in many cases, with limited impact on final quality.
Much less explored, however, has been the effect of soil applications during the early stages of fruit development, when the transpiration rate—and therefore the flow of calcium to developing tissues—is higher.
This article aims to analyze the variables that determine the potential success of early soil calcium applications to improve fruit firmness and condition at harvest and postharvest, contributing to the technical and economic sustainability of the sector in a context of strong international competition.
How does calcium accumulate in the fruit?
Fruit plants absorb calcium from the soil solution in the form of Ca²+, whose entry into the root occurs mainly through mass flow associated with water movement. Once in the rhizosphere, calcium can be mobilized to the xylem through the apoplastic, symplastic, or both pathways.
However, the preferential transport of calcium takes place through the apoplast, whose flow depends directly on the transpiration of the aerial parts of the plants (White, 2001).
Within the tree, calcium moves almost exclusively through the xylem and, consequently, its accumulation in different organs is determined by their transpiration intensity.
This, in turn, depends on the leaf area and stomatal density of each organ (Winkler and Knoche, 2019).
As a result, leaves accumulate higher Ca concentrations than fruits during the season, due to their greater transpiration.
However, during fruit development there is a critical window for the accumulation of this cation, which for cherries is located between fruit set and approximately 15–20 days after full bloom (DAFB) (Winkler et al., 2020); 30 DAFB for kiwifruit (Montanaro et al., 2015); 40 DAFB for avocado (White, 1990); and 25 DAFB for apricot (Montanaro et al., 2010), a period when fruit transpiration reaches its maximum (Figure 1).
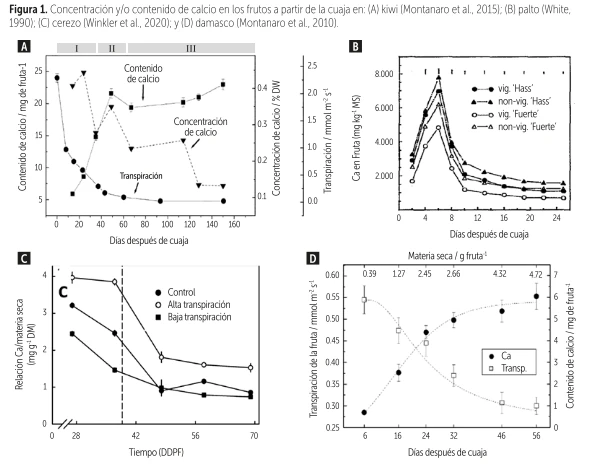 Figure 1. Calcium concentration and/or content in fruits from fruit set onwards in: (A) kiwifruit (Montanaro et al., 2015); (B) avocado (White, 1990); (C) cherry (Winkler et al., 2020); and (D) apricot (Montanaro et al., 2010).
Figure 1. Calcium concentration and/or content in fruits from fruit set onwards in: (A) kiwifruit (Montanaro et al., 2015); (B) avocado (White, 1990); (C) cherry (Winkler et al., 2020); and (D) apricot (Montanaro et al., 2010).
Firm fruit from the roots
The importance of early soil calcium application
The Chilean cherry is going through its most challenging moment: after a decade of expansion, the last season was marked by a decline in selling prices. Today, the industry’s competitiveness requires fruit of good size, firm, in excellent condition, and with maximum use of available resources.
In this context, calcium is a strategic ally in the fruit’s physiology, being decisive for firmness and postharvest life.
The importance of early soil calcium application
Over the last decade, the cherry tree has become the most relevant fruit crop in Chilean horticulture, reaching more than 76,000 hectares planted (ODEPA, 2025) and with a strong dependence on exports to China, which concentrates more than 90% of shipments (Muñoz, 2025).
However, in the last season, grower returns experienced a sharp drop due to relative oversupply and the increasing demand from the market for high-quality fruit.
This is a scenario where it becomes essential to improve efficiency in the use of resources while ensuring quality attributes that allow fruit differentiation and sustain competitive prices.
Among the main attributes, flesh firmness is critical, both for consumer preference and for its close relationship with resistance to mechanical damage, tolerance to long transport, and the incidence of postharvest rots.
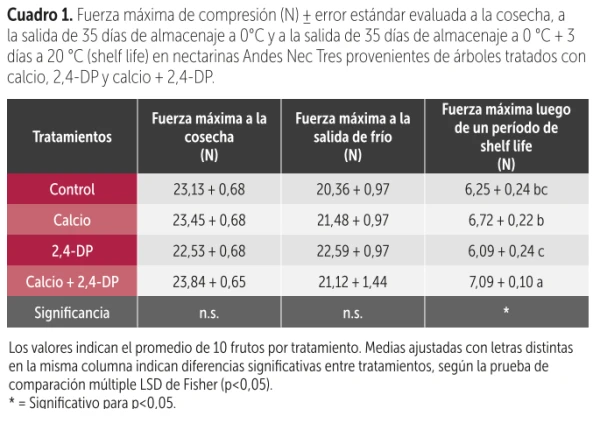 Table 1. Maximum compression force (N) ± standard error evaluated at harvest, after 35 days of storage at 0 °C and after 35 days of storage at 0 °C + 3 days at 20 °C (shelf life) in Andes Nec Tres nectarines from trees treated with calcium, 2,4-DP, and calcium + 2,4-DP.
Table 1. Maximum compression force (N) ± standard error evaluated at harvest, after 35 days of storage at 0 °C and after 35 days of storage at 0 °C + 3 days at 20 °C (shelf life) in Andes Nec Tres nectarines from trees treated with calcium, 2,4-DP, and calcium + 2,4-DP.
Role of calcium in firmness
Fruit firmness is largely determined by the structure and dynamics of the cell wall, where calcium plays an essential role.
This cation not only participates in key physiological processes such as signaling and membrane integrity but also stabilizes pectins in the middle lamella through the formation of “egg-box” bonds, which confer rigidity and reduce the loss of cellular cohesion.
Consequently, an adequate accumulation of calcium in the fruit can delay softening, extend shelf life, and increase the share of exportable fruit.
Although numerous studies have been conducted on foliar applications or postharvest dips with calcium-rich solutions, results have been inconsistent and often with limited impact on final quality.
Early stages and availability
Much less explored, however, has been the effect of soil applications during the early stages of fruit development, when the transpiration rate—and therefore the flow of calcium to developing tissues—is higher.
This article aims to analyze the variables that define the potential success of early soil calcium applications to improve fruit firmness and condition at harvest and postharvest, contributing to the technical and economic sustainability of the industry in a context of high international competition.
How does calcium accumulate in the fruit?
Fruit plants absorb calcium from the soil solution in the form of Ca²+, whose entry into the root occurs mainly through mass flow associated with water movement. Once in the rhizosphere, calcium can move into the xylem through the apoplastic, symplastic, or both pathways.
However, the preferential transport of calcium takes place through the apoplast, whose flow depends directly on the transpiration of the aerial parts of the plants (White, 2001).
Within the tree, calcium moves almost exclusively through the xylem and, consequently, its accumulation in different organs is determined by their transpiration intensity. This, in turn, depends on the leaf area and stomatal density of each organ (Winkler and Knoche, 2019).
As a result, leaves accumulate higher Ca concentrations than fruits during the season, due to their higher transpiration.
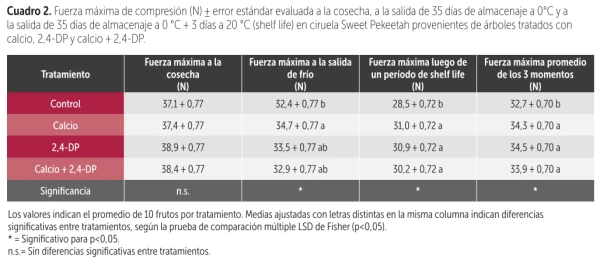 Table 2. Maximum compression force (N) ± standard error evaluated at harvest, after 35 days of storage at 0 °C and after 35 days of storage at 0 °C + 3 days at 20 °C (shelf life) in Sweet Pekeetah plums from trees treated with calcium, 2,4-DP, and calcium + 2,4-DP.
Table 2. Maximum compression force (N) ± standard error evaluated at harvest, after 35 days of storage at 0 °C and after 35 days of storage at 0 °C + 3 days at 20 °C (shelf life) in Sweet Pekeetah plums from trees treated with calcium, 2,4-DP, and calcium + 2,4-DP.
However, during fruit development there is a critical window for the accumulation of this cation, which for cherries occurs between fruit set and approximately 15–20 days after full bloom (DAFB) (Winkler et al., 2020); 30 DAFB for kiwifruit (Montanaro et al., 2015); 40 DAFB for avocado (White, 1990); and 25 DAFB for apricot (Montanaro et al., 2010), a period when fruit transpiration reaches its peak (Figure 1).
Once fruits surpass this critical stage, xylem functionality decreases drastically and the dilution effect reduces Ca concentration, limiting the effective accumulation window to just a few weeks, depending on the species and the growing region.
Indeed, in a study on apricot by Montanaro et al. (2010), results showed that over 83% of the total calcium content in the fruit accumulated during the first four weeks after fruit set, with the highest rate of accumulation in the first two weeks.
Furthermore, calcium uptake ceased as transpiration declined.
Environmental factors and management
This study concluded that fruit transpiration accounts for more than 60% of the total calcium entering the fruit during the season, and suggests that an optimal calcium availability in the soil and its distribution to the fruits during the first weeks of growth are essential to promote nutrient accumulation in the fruit.
Conversely, in areas with cold springs, high relative humidity, and prolonged cloudy skies, fruit transpiration decreases and the likelihood of obtaining soft fruit increases. A similar effect may occur with protective covers during the first weeks after fruit set, as these tend to increase ambient humidity and reduce fruit transpiration during the critical calcium accumulation period.
Therefore, calcium management in cherries requires a precise integration of early fertilization, understanding of fruit physiology, and adaptation to environmental conditions, with the goal of producing firm, high-quality fruit suitable for export.
Strategies to increase firmness
From an agronomic standpoint, early-season soil calcium application depends directly on the availability of this cation to stabilize pectins and provide structural rigidity. The synthesis and transport of auxins, particularly indole-3-acetic acid (IAA), are high during this period and may act as a physiological signal promoting calcium mobilization to fruits and other actively developing organs.
Based on this physiological rationale, a trial was conducted on Andes Nec 3 nectarines, a variety with a tendency to soften.
The treatment combined the application of 20 units of calcium (10 pre-bloom and 10 post-fruit set) together with the synthetic auxin 2,4-DP (75 mg L⁻¹), applied at 80% petal fall.
The results indicated that the combination of both strategies significantly improved fruit firmness (Table 1) and reduced mealiness after cold storage plus three days at 22 °C (Figure 1) (Quintanilla et al., 2025).
To validate the consistency of these results, the same treatments were applied on Sweet Pekeetah plums during the 2023–2024 season, where calcium, the growth regulator, and their combination all increased firmness compared to the control (Table 2).
These findings not only confirm the importance of an adequate calcium supply during early stages, but also highlight the potential of auxins as a physiological tool to direct and enhance its partitioning towards fruits.
However, further studies on other fruit species are needed to better understand the physiological mechanisms underlying these results and to optimize management of this technique for orchard implementation.
Is it necessary to apply soil calcium given that central Chile’s soils and waters are rich in this ion?
A frequent question in the industry is whether it is really necessary to apply calcium to the soil to improve fruit firmness, considering three main arguments:
- most soils in central Chile have high calcium concentrations,
- many irrigation waters also contain significant amounts of this nutrient,
- fruit accumulates, in absolute terms, relatively little calcium compared with other cations such as K⁺ or Mg²⁺.
These arguments are not unfounded. However, empirical and scientific evidence suggests an affirmative answer: yes, a targeted calcium fertilization strategy specifically for the fruit is necessary.
The rationale is both physiological and agronomic. During a very narrow period—from fruit set until cherry fruits reach 5–8 mm in diameter—it is essential to generate xylem sap extraordinarily rich in calcium, capable of supporting the intense processes of cell division and cell wall deposition that determine cherry firmness.
Under natural conditions, the calcium available in the soil may be sufficient to meet the tree’s basic physiological needs, but it is probably not enough to build the “super fruit” demanded by modern horticulture.
Obtaining “super fruit”
A cherry capable of maintaining its firmness, resisting a long journey to distant markets such as China, and being displayed on supermarket shelves with a quality standard equal to or greater than fruit produced closer to consumption centers.
In this context, soil calcium fertilization should not be seen merely as a nutrient supply, but as a strategic tool to modify calcium concentration in xylem sap during the critical accumulation window.
This approach, based on the physiology of mineral transport, allows growers to go beyond the “natural” conditions of tree nutrition and to direct its metabolism towards producing firmer fruits with longer postharvest life—indispensable attributes in today’s competitive scenario.
But are 20 or 30 kg of calcium enough?
To answer, consider this exercise: if a soil in central Chile has 10 meq/100 g of exchangeable calcium (a concentration classified as high) and a bulk density of 1.4 g/cm³, it should contain about 2.8 g/L of exchangeable calcium.
Assuming the roots of a tree actively colonize about 100 L of soil, each tree has access to about 280 g of exchangeable calcium in the portion of soil explored by its roots.
However, although the soil may hold a large calcium reserve, only a small fraction is actually dissolved in the soil solution and available to the roots at any given time. According to Mengel and Kirkby (2001), this amount is generally between 20 and 100 mg per liter of water, even in calcium-rich soils.
In other words, most calcium remains “stored” in the soil exchange complex and is released gradually as roots absorb it.
As Weil and Brady (2017) explain, there is a dynamic equilibrium: when the solution is depleted, it is replenished from the exchange reserve.
In simple terms, Sparks (2003) notes that in most agricultural soils, only about 1% of total exchangeable calcium is actually in solution at any given time.
Continuous calcium flow
This implies that the plant never accesses all the calcium at once but receives it through a continuous flow that depends on soil moisture, electrical conductivity, and the soil’s ability to replenish the solution.
An application of 30 kg/ha of elemental calcium to the soil is equivalent to about 30 g of calcium per tree in an orchard with 1,000 trees/ha.
Based on the concepts just described, and considering that the goal of soil calcium applications is to increase the concentration of calcium in the circulating solution, such an amount can significantly raise the natural concentration of available calcium, even in calcium-rich soils.
Early calcium applications
Role of vapor pressure deficit, soil temperature, and spring transpiration
In cherry production, another recurring topic is the efficiency of soil calcium applications carried out between September and October. Some agronomists argue that, at that time, low soil temperatures limit root activity and therefore calcium uptake, making fertilization ineffective.
However, plant physiology indicates that one of the main drivers of calcium transport from roots to developing fruits is not only soil temperature, but the air vapor pressure deficit (VPD), that is, the difference in vapor pressure between the evaporating surface (flowers, young fruits) and the surrounding air.
VPD is crucial because it strongly regulates transpiration and thus the water flow that is the main vehicle of calcium to the fruit.
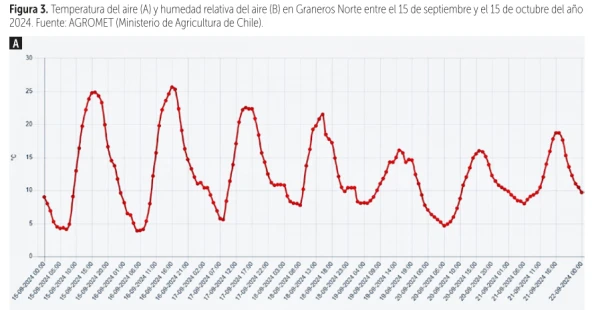
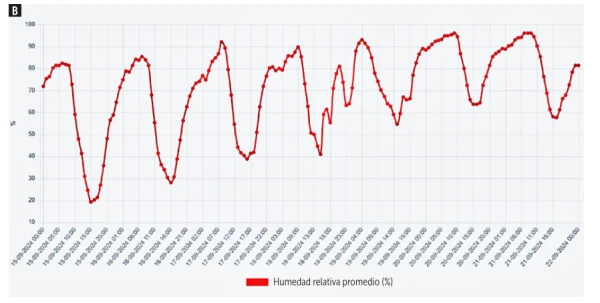 Figure 3. Air temperature (A) and relative humidity (B) in Graneros Norte between September 15 and October 15, 2024. Source: AGROMET (Ministry of Agriculture, Chile).
Figure 3. Air temperature (A) and relative humidity (B) in Graneros Norte between September 15 and October 15, 2024. Source: AGROMET (Ministry of Agriculture, Chile).
VPD classification
- Optimal VPD: 0.8 – 1.5 kPa. Enables sustained transpiration flow, favoring calcium arrival to developing fruits.
- Very low VPD: < 0.5 kPa. Indicates humid atmosphere; transpiration is limited and calcium mobilization is reduced.
- Very high VPD: > 1.8 kPa. Indicates very dry air; causes stomatal closure and limits both transpiration and nutrient transport.
To illustrate the practical relevance of this phenomenon, a concrete example can be taken from the Graneros area, O’Higgins region, Chile.
According to AGROMET (Ministry of Agriculture, Chile), on September 15, 2024, at 1:30 pm the air temperature was 22.2 °C and relative humidity was 30% (Figure 3).
Projected on a psychrometric chart, these data yield a VPD of 1.4 kPa, a value within the optimal transpiration range.
Spring transpiration and calcium uptake
This demonstrates that during early spring—at flowering, fruit set, and post-fruit set stages—atmospheric conditions can be sufficiently favorable for small fruits to undergo effective transpiration, mobilizing calcium into their tissues.
However, it is necessary to evaluate these variables in each production area to objectively understand whether trees are actually transpiring at a given time.
Another frequently raised objection regarding early calcium applications in fruit trees is the inefficiency of this practice in spring, since between September and October the soil may still be too cold to favor active root growth and thus adequate calcium uptake.
However, instrumental records show that this perception does not necessarily match reality. According to INIA’s database (2024) for Chimbarongo—a representative cherry production area in Chile—between September 15, 2024, and October 15, 2025, soil temperatures below 10 cm recorded minimum values ranging between 10 and 12 °C, while the average remained constantly around 15 °C, and at several points maximums exceeded 20 °C (Figure 4).
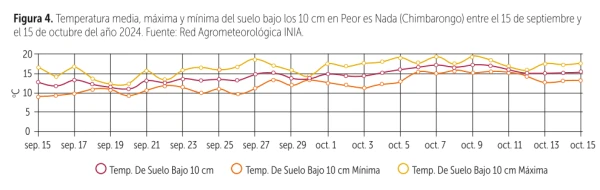 Figure 4. Average, maximum, and minimum soil temperature at 10 cm depth in Peor es Nada (Chimbarongo) between September 15 and October 15, 2024. Source: INIA Agrometeorological Network.
Figure 4. Average, maximum, and minimum soil temperature at 10 cm depth in Peor es Nada (Chimbarongo) between September 15 and October 15, 2024. Source: INIA Agrometeorological Network.
Soil temperature and root activity
These ranges are compatible with the metabolic reactivation of the roots and with the processes of absorption of essential nutrients, including calcium.
In addition, according to the same analysis, below 40 cm the soil temperature remains stable at 15 °C throughout the year, providing a uniform environment for the functioning of the deeper root system.
Consequently, and in general terms, soils in the central zone at this phenological stage would not present such extreme limiting conditions as is often assumed, but rather offer an adequate context for early calcium applications to contribute to nutrient replenishment in growing tissues, where its structural incorporation is critical for fruit development and quality.
Here too, it is necessary to evaluate each production area with objective data to verify the thermal conditions in which roots are found in spring.
And what about calcium reserves?
A little-discussed aspect in fruit tree calcium nutrition is the role of calcium reserves present in fruitwood and perennial structures.
Some experimental evidence shows that the calcium concentration in the bark decreases progressively from budbreak onwards (Mason and Whitfield, 1960), while at the same time a high calcium concentration in the xylem vessels is observed at the start of the growing season (Wieneke, 1969; Ferguson and Turner, 1981).
This behavior is particularly logical in deciduous species that flower before producing new shoots, since the initial calcium demand could not be met solely by early root uptake.
It is now known that calcium can be stored in the form of calcium oxalate (CaOx) in the wood, derived from ascorbate synthesis, and that there are specific enzymes responsible for the degradation of these crystals when the nutrient is again required for metabolic functions, such as the enzyme oxalate oxidase.
Distribution and function of CaOx
However, some relevant questions remain open: where is this CaOx mainly located that could supply fruits during the early stages? A plausible hypothesis is that it is found in the fruitwood — spurs, shoots, and main branches — structures that play a key role as reserve and support organs.
Regarding its formation, it is hypothesized that CaOx accumulates in conjunction with transpiration processes: calcium, transported in the xylem flow, would precipitate as oxalate in tissues characterized by high stomatal activity.
Based on this hypothesis, the critical moment for the accumulation of such reserves would be during the differentiation of spurs and shoots, while these structures are still green and with functional stomata.
Once lignified, transpiration decreases drastically and the capacity to further accumulate calcium in the form of oxalate would be limited.
These questions, which are fundamental for designing a calcium nutrition management strategy aimed at enhancing calcium accumulation in reserve organs, are currently being studied by our research group, with the aim of soon providing new information to improve calcium fertilization practices in orchards.
Opening image source: SL Fruit Service
José Ignacio Covarrubias, Fernanda Vasquez, Francisco Quintanilla and Thomas Fichet
Redagricola
Cherry Times - All rights reserved








